Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Результаты генетических и эмбриологических исследований гомеозисных генных комплексов Antennapedia и bithorax показывают, что эти локусы действительно играют роль переключателей, определяющих судьбу отдельных сегментов. Переключение происходит в период детерминации и, по-видимому, представляет собой результат позитивного действия названных локусов этих комплексов на батареи других генов. Следует отметить, что альтернативный путь развития, происходящий в отсутствие этих генов, не бывает хаотическим; в норме он наблюдается у некоторых других частей животного, так что эти гены можно в самом деле рассматривать как переключатели, производящие выбор из нескольких альтернативных состояний. Изменение реакции, вызванное гомеозисной мутацией, не приводит, как хотелось бы Гольдшмидту, к появлению «перспективных монстров», обладающих потенциальными возможностями для макроэволюционных событий. На самом деле изменения эти носят атавистический характер и могут дать сведения об истории генетических регуляторных механизмов, приобретенных в процессе эволюции. Вероятно, особенно ясно это можно продемонстрировать на примере эволюционной истории насекомых.
Филогения членистоногих, или как гомеология повторяет филогению
Филогенетические взаимоотношения крупных групп членистоногих представлены на рис. 8-10. Эта филогения построена на основании данных палеонтологических, эмбриологических, сравнительно-анатомических и различных других исследований.
Членистоногие возникли от некого предка, сходного с кольчецами, по всей вероятности, в конце докембрия, а первая значительная радиация этой группы началась в кембрии. Среди ископаемых остатков, найденных в нижнепалеозойских отложениях, обнаружены представители всех групп членистоногих, за исключением насекомых. Насекомые появляются в верхнем палеозое, т. е. примерно 350 млн. лет назад. Если в отношении происхождения членистоногих от кольчецов (или от какой-то сходной с ними группы) существует известное единодушие, то по вопросу о том, являются ли главные группы членистоногих моно- или полифилетическими, единого мнения нет. Мы не собираемся вступать в эту дискуссию, а сконцентрируем внимание на той части филогении, которая относится к группе членистоногих Uniramia и от которой в конечном счете произошли насекомые. Изменения в характере сегментации у этой группы членистоногих можно проанализировать в свете гомеозисных мутаций дрозофилы.
К признакам, позволяющим считать кольчецов предками членистоногих, относятся лежащая в основе их строения метаметрия, расположение нервной системы на вентральной стороне, а сердца - на дорсальной стороне тела; это признаки, которыми обладают все членистоногие. Несмотря на эти общие для обеих групп признаки взрослых особей, они сильно различаются по ранним стадиям развития. Для кольчецов, так же как для моллюсков, характерно спиральное дробление, тогда как у членистоногих дробление в общем центролецитальное, описанное выше для дрозофилы. Поэтому одним из крупнейших событий в возникновении членистоногих было резкое изменение раннего развития при сохранении взрослыми особями основного плана строения.
На рис. 8-11 показаны поздние стадии развития морской полихеты Polydora ciliata. После дробления и гаструляции у этого червя образуется свободноплавающая личинка - трохофора. Эта личинка разделена двумя венчиками ресничек на три обособленные области. Передняя, предротовая, часть отделена от средней, туловищной, части прототрохом. Задняя, пигидиальная, отделена от средней части телотрохом. В процессе развития личинки ее туловищная часть разделяется на три обособленных туловищных сегмента. На этой стадии у личинки можно различить в общей сложности шесть сегментов: предротовой, ротовой, три туловищных и хвостовой, или пигидий. Личинка продолжает расти путем добавления новых сегментов в зоне нарастания, находящейся между последним туловищным сегментом и пигидием. Этот рост кзади сопровождается постепенным усложнением структур, специфичных для каждого вида, на самых задних сегментах. После того как личинка достигнет полного развития, она в результате метаморфоза превращается во взрослого червя. Этот последний вопрос нас здесь не интересует; нам достаточно отметить сегментарную структуру животного и то, как эта сегментарная структура изменяется на «следующей» стадии филогенеза насекомых.
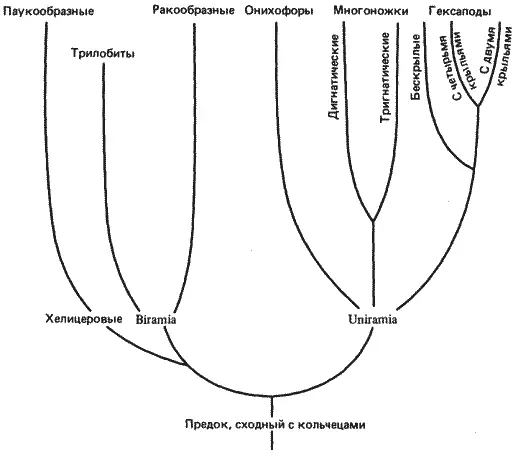
Рис. 8-10.Филогенетические связи между главными группами членистоногих с упором на линию, давшую начало современным насекомым (Hexapoda).
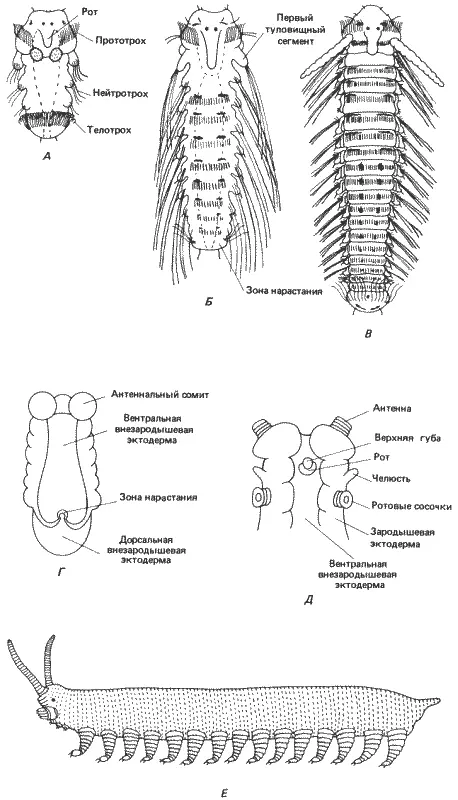
Рис. 8-11. А-В. Сегментация тела морской полихеты Polidora. А. Ранняя личиночная стадия. Б. Более поздняя личиночная стадия после добавления нескольких сегментов к зоне нарастания на заднем конце тела. В. Молодая взрослая особь. Г-Е. Зародыши и взрослая форма онихофор. Г. Вентральная сторона зародыша Peripatoides novaezealandiae на ранней стадии образования зародышевой полоски; как и у кольчецов, добавление новых сегментов происходит на заднем конце тела. Д. Вентральная сторона головы зародыша P. capensis, состоящей из трех сегментов - антеннального, челюстного и ротового - и головных сосочков. Е. Обобщенная взрослая онихофора (рисунок сделан с зафиксированных экземпляров); все туловищные сегменты одинаковы, и на каждом из них имеется по одной паре членистых ходильных ножек с коготком на конце ( А-В - Wilson, 1928; Г - Sheldon, 1889; Д - Sedgwick, 1888).
Когда впервые были открыты онихофоры, их сочли «недостающим звеном» между кольчецами и высшими членистоногими. На первый взгляд это кажется правильным, и они в самом деле представляют собой промежуточную форму. Однако онихофоры не являются прямыми предками насекомых. Дробление у них центролецитальное, и у видов, имеющих крупные, богатые желтком яйца, бластодерма образуется на поверхности массы желтка путем целлюляризации. В процессе гаструляции на среднебрюшной поверхности бластодермы формируются две эктодермальные зародышевые полоски и происходит инвазия мезодермальных клеток в бластодерму. Крупные гаструляционные перемещения клеток возникают только в связи с образованием переднего зачатка средней кишки. Эти перемещения начинаются на презумптивном переднем конце животного и распространяются к хвостовому его концу. Вслед за началом этих перемещений начинается процесс сегментации. Ранняя стадия сегментации Peripatoides novaezealandiae показана на рис. 8-11, Г . На переднем конце животного имеется хорошо заметный антеннальный сегмент, за которым на этой стадии следует ряд сходных сегментов, дающих сомиты. Как и у личиночных форм полихет, новые сегменты образуются субтерминально, в задней зоне нарастания. Сегментация завершается до рождения на свет (большинство видов онихофор живородящие или яйцеживородящие). На рис. 8-11, Д изображена более поздняя стадия эмбриогенеза Peripatopsis capensis, соответствующая началу развития головы. Голова состоит из переднего антеннального сегмента, единственного челюстного сегмента, на котором находится рот, и сегмента, несущего ротовые сосочки, где открываются протоки слизистых желез. Позади головы расположен ряд одинаковых туловищных сегментов, от каждого из которых у взрослой особи отходит членистая нога, снабженная коготком.
Читать дальшеИнтервал:
Закладка:









