Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Вторая причина связана с тем, что мультигенные семейства должны обеспечивать переключение генов в процессе развития. Семейства, содержащие близкие, но не идентичные гены, продуцируют сходные продукты, каждый из которых специфически необходим или клеткам различных типов, или на разных стадиях развития. Наиболее хорошо изученным примером служат небольшие мультигенные семейства, содержащие глобиновые гены. Эволюционные связи между глобинами человека, установленные на основании их аминокислотных последовательностей, представлены на рис. 10-4. Предковый гемоглобин дивергировал от миоглобина примерно ко времени возникновения хордовых, в конце докембрия. В свою очередь предковый β-глобин дивергировал от предкового α-глобина примерно 500 · 10 6лет назад, на заре истории позвоночных, в раннем палеоцене. Фетальная γ-цепь дивергировала от β-цепи самое большее 200 · 10 6лет назад, в самый начальный период истории млекопитающих. Наконец, δ-цепь (вариант β-глобина), входящая в качестве минорного компонента в нормальный гемоглобин взрослого человека, дивергировала от β-цепи примерно 40 · 10 6лет назад. Гены α-цепей образуют небольшое мультигенное семейство из трех членов, а гены β-цепей - семейство из семи генов.
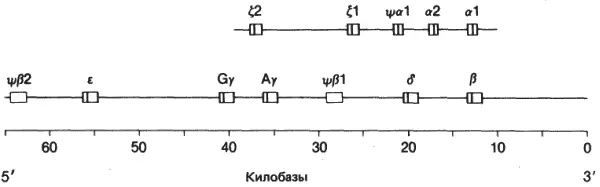
Рис. 10-4.Распределение генов α- и β-подобных глобинов человека. На шкале указана длина хромосомной ДНК в килобазах. Глобиновые гены изображены в виде прямоугольников. Черные поперечные полосы соответствуют кодирующим последовательностям, а белые участки-нитронам. Псевдогены (ψ) изображены в виде чисто белых прямоугольников. Транскрипция в направлении 5'–>3' идет слева направо (Efstratiadis, 1980).
Функциональная молекула гемоглобина - это тетрамер, состоящий из двух субъединиц α-типа и двух субъединиц β-типа, т.е. α 2β 2. У человека имеется 7 образующих кластер генов глобинов β-типа, расположенных так, как это показано на рис. 10-4. Два из них кодируют Aγ- и Gγ-цепи, обнаруженные в фетальном гемоглобине α 2γ 2; два гена, δ и β, кодируют цепи, экспрессируемые после рождения ребенка в преобладающих (α 2β 2) и минорных (α 2δ 2) видах гемоглобина. У человека имеется также другой глобин β-типа ε и глобин α-типа - ζ, которые экспрессируются только в эмбриональном гемоглобине ζ 2ε 2. Два других члена β-семейства, ψβ 1и ψβ 2, - неэкспрессирующиеся псевдогены. Расписание переключения синтеза с одного глобина на другой в процессе развития человека показывает изменение времени экспрессии генов глобина и мест, в которых они экспрессируются.
На ранних стадиях зародышевого развития ζ-и ε-цепи синтезируются содержащими ядро мегалобластами, образующимися в желточном мешке. Этот синтез быстро снижается и на шестой неделе развития заменяется синтезом другого типа, в котором безъядерные эритроциты, происходящие из стволовых клеток печени и селезенки, синтезируют α- и γ-глобиновые цепи, характерные для плода. На поздних стадиях развития плода костный мозг становится главным местом синтеза глобинов. Вскоре после рождения происходит вторичное переключение глобинового синтеза и переход к синтезу гемоглобина взрослого типа (adult). У этого переключения есть одна очень существенная особенность: переход от одного типа синтеза к другому связан с генной регуляцией в отдельных стволовых клетках, а не просто с заменой стволовых клеток, продуцирующих γ-глобин, на стволовые клетки, продуцирующие β-глобин, потому что во время переключения отдельные эритроциты вырабатывают как γ-, так и β-глобиновые цепи.
Переключение глобиновых генов - общее правило для всех позвоночных, однако типы переключения удивительно разнообразны даже среди млекопитающих. У человека имеются хорошо различимые эмбриональные, фетальные глобины и глобины взрослого типа. Фетальный гемоглобин обладает более высоким сродством к кислороду, чем материнский гемоглобин, что облегчает перенос кислорода через плаценту к плоду. Однако, как показано в табл. 10-5, не у всех млекопитающих имеются ясно выраженные фетальные гемоглобины. У грызунов, хищных и лошадей, например, на стадии плода происходит прямой переход от синтеза эмбриональных глобинов к глобинам взрослого типа. В этих случаях сродство к кислороду гемоглобина взрослого типа, находящегося в эритроцитах плода, по-видимому, как-то модулируется содержащимися в цитоплазме малыми молекулами, так что кровь плода обладает более высоким сродством к кислороду, чем материнская кровь.
Организация генов β-глобинов может быть очень сходной даже у организмов, у которых обнаружены существенные различия в типах переключения. Как видно из табл. 10-5, у кроликов синтезируются эмбриональные β-подобные глобины, называемые ε(Υ) и ε(Ζ), но у них нет фетального β-глобина, эквивалентного γ-цепям человека. Вместо этого плод синтезирует β-цепи взрослого типа. Лейси (Lacy) и ее сотрудники, а также Хардисон и др. (Hardison et al.) установили, что гены β-глобина у кролика образуют кластер, очень сходный с кластерами, имеющимися у человека, и что в процессе развития происходит переключение отдельных генов этого кластера. Два гена, соответствующие по своему местоположению γ-генам в кластере β-генов человека, показанном на рис. 10-4, экспрессируются у зародыша, вероятно для того, чтобы продуцировать ε-цепи. Ген, соответствующий по своему положению δ-гену человека, по-видимому, не экспрессируется ни на одной стадии развития, а ген, соответствующий гену β-глобина взрослого человека, выполняет ту же функцию у кролика.
Таблица 10-5. Переключения генов гемоглобина у некоторых млекопитающих
(по данным Hardison et al., 1979; Kitchen, 1974; Kitchen, Brett, 1974)
| Стадия развития | Кролик | Овца | Лошадь | Медвежий макак | Человек |
|---|---|---|---|---|---|
| Зародыш | λ 2ε(у) 2 | α 2ε 2 | Sα 2ε 2 | Отсутствуют | ζ 2ε 2 |
| χ 2ε(z) 2 | Fα 2ε 2 | ||||
| Плод | Отсутствуют | α 2γ 2 | Отсутствуют | α 1 2γ | α 2 Aγ 2 |
| α 2 2γ | α 2 Gγ 2 | ||||
| Взрослая особь | α 2β 2 | α 2β 2 | Sα 2β 2 | 1α 2β 2 | α 2β 2 |
| Fα 2β 2 | 2α 2β 2 | α 2δ 2 |
Глобины иллюстрируют роль переключения в экспрессировании последовательного ряда генов, непосредственно участвующих в метаболических функциях; существуют, однако, и более сложные системы мультигенов, продукты которых непосредственно участвуют в экспрессии генов или в морфогенезе. В некоторые из таких семейств входит большое число генов, экспрессия которых регулируется в ходе развития.
Гистоновые гены, лучше всего изученные у морских ежей, хорошо иллюстрируют адаптацию, состоящую в использовании мультигенных семейств для обеспечения как синтеза больших количеств белка, необходимых на ранних стадиях развития, так и переключений, необходимых для последовательной экспрессии генов ряда близких белков. В процессе дробления число ядер, а тем самым и содержащихся в них хромосом, быстро растет, увеличиваясь вдвое за каждые 10-20 мин. Гистоны, необходимые для обеспечения сборки хромосом во время быстрого дробления, могут поступать либо из белков, запасенных в яйце, как это показали Вудленд и Адамсон (Woodland, Adamson) на лягушке Xenopus, либо за счет усиленного синтеза гистонов в дробящемся зародыше, как установили Кедес (Kedes) и его сотрудники для морских ежей. Количество гистоновых генов, необходимых для этих двух стратегий, различно. В яйце лягушки гистоны медленно накапливаются в течение нескольких недель или месяцев оогенеза, тогда как в зародыше морского ежа они синтезируются за несколько часов. Яйцо морского ежа содержит лишь около 25% гистоновой мРНК, необходимой для синтеза гистонов при дроблении; остальная мРНК транскрибируется в процессе дробления. Вследствие этого главное семейство гистоновых генов представлено у морского ежа гораздо большим числом копий, чем у лягушки (или у человека). У морских ежей число копий этих генов колеблется от 300 до 1200, тогда как у Xenopus и у человека, по данным Бирнстила и др. (Birnstiel) и Вилсона (Wilson) и его сотрудников, их всего 10-20.
Читать дальшеИнтервал:
Закладка:









