Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Как установлено, у морских ежей гистоновый синтез регулируется сложным комплексом переключений, что приводит как к временным, так и к тканеспецифичным изменениям. Ньюрок (Newrock) и его сотрудники описали во всех подробностях сложную последовательность этих изменений, впервые обнаруженных Рудерманом и Гроссом (Ruderman, Gross). Из пяти главных гистонов морских ежей три (H1, H2A и Н2В) представлены на ранних стадиях дробления специфичными для этих стадий подтипами, образующимися в результате кратковременного синтеза. Далее происходит синтез гистонов других подтипов, специфичных для отдельных стадий и последовательно сменяющих друг друга, как это показано на рис. 10-5. Подтипы каждого гистона, специфичные для дробления, сменяются в процессе дробления подтипами α-гистонов, а затем на стадии бластулы β-, γ- и другими подтипами. Эксперименты Ньюрока и его сотрудников, Кункеля и Вейнберга (Kunkel, Weinberg) и Чайлдса и др. (Childs et al.) показывают, что замены гистоновых белков происходят в результате постепенных и последовательных изменений в синтезе гистоновой мРНК. Каждый подтип - продукт отдельного гена, относящегося к тому же семейству, что и гены, кодирующие другие подтипы. Замена гистонов одного подтипа гистонами другого подтипа приводит к изменению белкового состава хромосом по мере развития. Такие изменения могут привести к «ремоделированию» хроматина, что имеет потенциально важное значение для дифференцировки клеток зародыша.
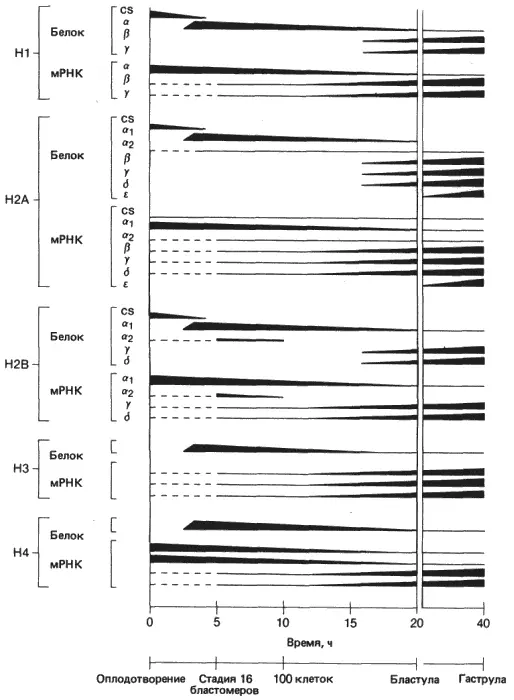
Рис. 10-5.Экспрессия членов гистоновых мультигенных семейств в процессе развития зародыша морского ежа. Толщина поперечных полос схематически отражает уровень синтеза белка или количество присутствующей мРНК. Обратите внимание, что синтез гистонов контролируется путем регуляции как трансляции, так и синтеза мРНК, потому что, хотя запасенные в яйце мРНК имеются в наличии на ранних стадиях развития, немедленной их трансляции не происходит (Newrock et al., 1977; Childs, Maxson, Kedes, 1977-с изменениями; использованы дополнительные данные для определения сроков начала трансляции гистонов на запасенной мРНК: Wells et al., 1981, а также Wells, Cohen, неопубликованные данные).
Переключения не ограничиваются такими белками, как глобины и гистоны, участвующие главным образом в обеспечении внутренних потребностей клетки. Некоторые переключения тесно связаны с морфогенезом. Например, микротрубочки, которым принадлежит ведущая роль в клеточном движении и создании формы клетки, состоят из α- и β-тубулинов, синтез которых обеспечивается небольшими мультигенными семействами. Э. Рэфф (Е. Raff) и ее сотрудники установили, что синтез этих тубулинов в процессе развития дрозофилы регулируется путем переключений. Некоторые виды тубулинов синтезируются на протяжении всего развития, но синтез по меньшей мере одного β-тубулина включается, а затем выключается в течение короткого периода эмбриогенеза. Кемпхьюз и др. (Kemphues et al.) обнаружили существо вание тканеспецифичного β-тубулина, экспрессирующегося только в семенниках. Этот специфичный для семенников β-тубулин необходим для сборки очень специализированной структуры - аксонемы хвоста сперматозоида.
Кафатос (Kafatos) и его сотрудники исследовали роль переключателей мультигенов в морфогенезе на примере белков хориона, образующих оболочку яйца, у шелкопряда. Если рассматривать невзрачную на вид оболочку яйца при помощи сканирующего или обычного электронного микроскопа, то обнаруживается ее изящная поверхностная структура, которая, несомненно, понравилась бы Д'Арси Томпсону. На рис. 10-6 показана поверхность хориона шелкопряда Antheraea polyphemus. Главные особенности строения этой оболочки - выстилающие ее шестиугольники, соответствующие местам, где прежде находились фолликулярные клетки, и выступающие вверх дыхательные структуры, удачно названные аэропилями. Видное на переднем плане углубление - это микропиле, через которое при оплодотворении в яйцо проникает сперматозоид. На поперечном срезе можно видеть, что хорион обладает легкой, но механически прочной структурой, укрепленной внутренними перекладинами. Хорион секретируется слоем фолликулярных клеток, синтезирующих белки; число различных белков хориона, синтезируемых у шелкопряда, приближается к 200. Эти белки можно разбить на пять больших классов, различающихся по молекулярной массе и последовательностям аминокислот. Белки, входящие в каждый класс, кодируются семейством близких генов, но, как показали Джонс и др. (Jones et al.), в пределах надсемейства белков хориона обнаружена, кроме того, значительная близость между ограниченными участками, или доменами, белков, принадлежащих к различным семействам. На близость генов, кодирующих эти белки, указывают также данные Мэрией Гольдсмит (Marian Goldsmith) и ее сотрудников о том, что у тутового шелкопряда Bombyx mori многие гены, кодирующие белки, относящиеся к этим семействам, организованы в три кластера, локализованные в одной и той же хромосоме.
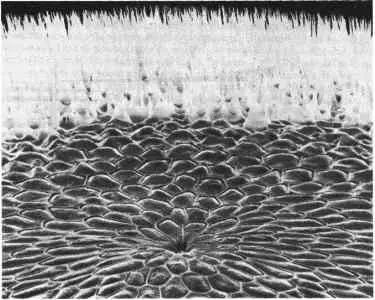
Рис. 10-6.Структура поверхности оболочки яйца шелкопряда Antheraea polyphemus. На переднем плане микропиле с концентрическими рядами выстилающих клеток вокруг, а на заднем плане - многочисленные аэропиле (структуры, обеспечивающие дыхание) (Regier, Mazur, Kafatos, 1980; в этой работе дано подробное описание хориона).
Экспрессия генов, кодирующих белки хориона, регулируется рядом переключений. Сим и др. (Sim et al.) установили, что на ранних стадиях развития хориона преобладает синтез членов трех семейств белков с высокой молекулярной массой, а на средних и поздних стадиях-синтез белков, относящихся к двум семействам с низкой молекулярной массой. При помощи соответствующих проб, состоящих из клонов рекомбинантной ДНК генов хориона, Симу и др. удалось выявить виды мРНК, соответствующие клонированным генам. Изменяющийся спектр синтезируемого белка отражает изменения спектра синтезируемой мРНК в фолликулярных клетках.
Два наблюдения, сделанные недавно в лаборатории Кафатоса, убеждают в том, что гены, детерминирующие развитие хориона, близки не только эволюционно и функционально; они, кроме того, расположены в определенном порядке, координирующем их экспрессию. У тутового шелкопряда Bombyx mori известна мутация, которая приводит к развитию дефектного хориона в результате делеции участка ДНК, содержащего примерно половину генов хориона. Утрачиваемые при этом гены относятся главным образом к тем, которые экспрессируются на поздних стадиях хориогенеза, позволяя предполагать, что гены собраны в кластеры в зависимости от сроков их экспрессии. Использование клонов ДНК, полученных путем рекомбинации фрагментов ДНК, которые содержат более одного гена, специфичного для хориона, дало возможность Джонсу и Кафатосу исследовать генную организацию с точки зрения временной регуляции экспрессии. Расположение генов в двух таких клонах показано на рис. 10-7. Эти два клона содержат различные гены, но в каждом клоне имеется по две копии этих генов. Гены эти принадлежат к разным подсемействам хорионных генов, но их экспрессия координирована. Гены, изображенные на рис. 10-7 в верхнем фрагменте, используются в середине хориогенеза, а гены нижнего фрагмента - в конце этого процесса. В обоих случаях физически смежные пары генов, по-видимому, остаются сцепленными при транскрибировании и между каждой парой находится общий для них регуляторный элемент.
Читать дальшеИнтервал:
Закладка:









