Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Таблица 6-1. Типы гетерохронии(Gould, 1977; с небольшими изменениями)
| Изменение сроков | Название по системе де Бера | Морфологический результат | |
|---|---|---|---|
| появления соматического признака | созревания репродуктивных органов | ||
| Ускорение | Без изменения | Акселерация | Рекапитуляция (путем ускорения) |
| Без изменения | Ускорение | Педоморфоз (прогенез) | Педоморфоз (путем урезывания) |
| Задержка | Без изменения | Неотения | Педоморфоз (путем задержки) |
| Без изменения | Задержка | Гиперморфоз | Рекапитуляция (путем продления) |
Сдвиги сроков развития могут привести к эволюционным результатам, прямо противоположным рекапитуляции - к педоморфозу. При педоморфозе признаки, которые у предков были характерны для ювенильных стадий, сохраняются у их потомков во взрослом состоянии; это, однако, может произойти двумя совершенно различными способами. В наиболее хорошо знакомом случае - при неотении - соматическое развитие задерживается по сравнению с течением полового созревания. Например, аксолотлю - этой рабочей лошадке неотении (рис. 6-1) - требуется столько же времени для полового созревания, как и родственным ему ненеотеническим хвостатым амфибиям, но при этом многие соматические морфологические признаки остаются у него на личиночной стадии, так что половозрелый аксолотль продолжает жить в воде и выглядит как личинка-переросток. Существует и другой вид педоморфоза - прогенез, который возникает в результате совершенно иного процесса. При прогенезе созревание гонад ускоряется, так что половая зрелость достигается в маленьком и в, сущности, ювенильном теле: соматическое развитие урезано.
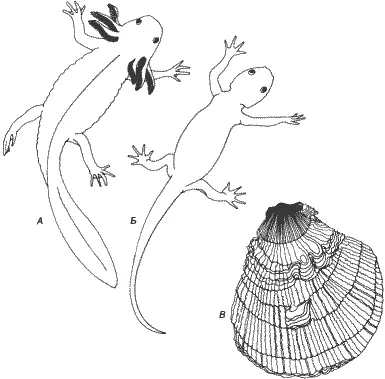
Рис. 6-1.Педоморфоз и рекапитуляция. А. Мексиканский аксолотль сохраняет жабры, уплощенный хвост и кожу, характерные для личинки, хотя он достигает размеров взрослой особи и половой зрелости. Если этой неотенической личинке вводить тироксин, то она претерпевает метаморфоз, превращаясь в типичную наземную взрослую форму (Б). В. У двустворчатого моллюска Hinnites наблюдается классическая рекапитуляция. На ювенильных стадиях Hinnites похож на других морских гребешков, но при дальнейшем созревании он прикрепляется ко дну и в результате продолжающегося роста раковины образует сходную с устрицей форму с менее упорядоченной структурой (Аксолотли с фотографий, любезно предоставленных G. M. Malacinski; Hinnites - с натуры).
Педоморфоз как одного, так и другого типа обеспечивает легкую в генетическом отношении эволюционную реакцию на давление со стороны среды, потому что при этом может быть использована морфогенетическая система, которая уже интегрирована. И неотения, и прогенез приводят к педоморфозу, но они, по-видимому, возникают в ответ на разные условия. Неотения может, например, возникнуть в ситуациях, когда неотеническая особь получает возможность избежать перехода в суровую или неустойчивую среду, в которой должен обитать взрослый организм, и остаться в более мягкой и стабильной среде, где обитают личинки. Типичным примером этого служат неотенические личинки амфибий; у некоторых групп хвостатых амфибий в процессе эволюции возникли неотенические взрослые формы, продолжающие жить в воде.
Неотения открывает довольно неожиданные эволюционные возможности, как, например, у нелетающих птиц, питающихся на земле. Неспособность к полету часто встречается у птиц, обитающих на островах, где отсутствуют хищные млекопитающие. Поскольку, как указывают Олсон (Olson) и Федуччиа (Feduccia), летательные мышцы, кости плечевого пояса и грудина составляют примерно 20-25% массы обычной птицы, то на их рост и поддержание затрачивается значительная доля ее энергетического бюджета. Если полет не дает непосредственного преимущества, то отбор не будет благоприятствовать сохранению дорогостоящего в энергетическом выражении комплекса структур. Одним из механизмов, обеспечивающих быструю утрату этих признаков, может быть неотения, и нелетающие птицы действительно во многом сходны с птенцами летающих видов. Согласно Олсону и Федуччиа, у некоторых групп, таких как голуби и пастушки, грудина развивается на поздних стадиях онтогенеза, после вылупления. Разница между ними и куриными значительна: среди куриных нет ни одного нелетающего вида. У типичного представителя куриных - курицы - окостенение грудины начинается между 8-м и 12-м днями инкубирования, а не после вылупления, как у пастушков. Задержка части морфогенетической программы на такой ранней стадии развития привела бы, возможно, к летальному исходу, однако задержка развития грудины у форм, у которых эти процессы происходят лишь после вылупления, вряд ли вызовет серьезные осложнения. Таким образом нелетающие пастушковые возникали в процессе эволюции неоднократно, потому что в программе развития пастушковых имеется нреадаптация к изменению такого типа. После того как неотеническое изменение возникло, становится возможным достижение больших размеров и другие адаптации. К сожалению, дронты, моа, эпиорнисы и все другие многочисленные виды, составлявшие великолепный ряд нелетающих птиц, населявших большие и малые острова по всему земному шару, вымерли в результате вмешательства человека в природу.
Неотенические признаки обычны у птиц и млекопитающих с высокоразвитым общественным образом жизни, и было высказано предположение, что задержанное соматическое созревание, возможно, облегчает распознавание ранга отдельных особей, позволяя избегать столкновений и повышать стабильность сообщества. Неотенические признаки человека отражают в усиленной степени такие признаки приматов, как стабильность сообщества, интенсивная забота о потомстве и длительный период созревания. В строении черепа отражены также такие Неотенические признаки, как увеличение объема головного мозга, которое было достигнуто путем увеличения продолжительности внутриутробного развития с сопровождающими его аллометрическими тенденциями.
В отличие от этого прогенез представляет собой, по-видимому, типичную реакцию на такие условия среды, в которых высокий уровень размножения или малые размеры отдельной особи могут оказаться особенно выгодными. В некоторых из таких случаев отбор может идти не на какой-то определенный морфологический признак, а на малые размеры как таковые. В результате возникает зрелый в репродуктивном отношении организм, обладающий личиночной морфологией или же смесью личиночных и взрослых признаков. В средах, открытых для небольших организмов, некоторые морфологические признаки выходят из-под ограничительных воздействий отбора, которые испытывали на себе более крупные предковые виды. В условиях ослабленного или модифицированного отбора анатомические признаки могут накапливать изменения, делающие возможным переходы от одного плана строения к другому, осуществить которые в других условиях было бы крайне трудно. Подобная точка зрения вполне приложима также к происхождению насекомых от педоморфных многоножек, как это предполагает де Бер. На рис. 6-2 изображена личинка многоножки Glomeris, у которой в момент вылупления имеется только три пары ног и ограниченное число сегментов тела. Как правило, число ног и сегментов тела увеличивается у многоножек в процессе развития. Если, однако, такая личинка достигла бы половозрелости, то получился бы организм, очень похожий на примитивное насекомое.
Читать дальшеИнтервал:
Закладка:









