Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
У домовой мыши (Mus musculus) имеется один сложный ген, Т -локус, который во многом сходен с только что описанной системой Notch. Первый аллель этого локуса был описан как аутосомный доминантный аллель, названный Brachyury (Т). У мышей, гетерозиготных по этому гену, Т/+, хвосты короткие. В гомозиготном состоянии (Т/Т) этот ген летален и зародыши гибнут внутриутробно. Вскоре после обнаружения этой доминантной мутации было установлено, что потомки от скрещивания гетерозигот ( Т/+ ) с мышами дикого типа часто вовсе лишены хвоста. Оказалось, что эти бесхвостые мыши - результат рецессивных аллелей Т -локуса, часто встречающихся в природных популяциях мышей. Следовательно, эти бесхвостые мыши имели генотип T/t. От скрещиваний между такими гетерозиготными мышами T/t были получены бесхвостые мыши, размножающиеся в чистоте. Позднее было показано, что это обусловлено «сбалансированной системой леталей». Как оказалось, среди потомков от скрещивания гетерозигот летальными были не только гомозиготы Т/Т, чего следовало ожидать, но и гомозиготы t/t. Таким образом, выживали только гетерозиготы T/t, которые и давали следующее поколение. Эта интригующая ситуация была изучена как генетически, так и эмбриологически в ряде изящных работ Денна (Dunn), его учеников Беннета (Bennett) и Глюксон-Вэлша (Gluechsohn-Waelsch).
В результате генетических исследований вновь полученных линий, сбалансированных по леталям, было установлено, что скрещивания между бесхвостыми мышами, получившими свой рецессивный аллель t от разных популяций, часто давали нормальных потомков. В частности, от скрещивания T/t a x T/t b были получены нормальные и бесхвостые потомки в соотношении 2:1. Можно показать, что потомки с нормальными хвостами имеют генотип t a/t b . Таким образом, этот генотип был не только нелетальным, но и нормальным морфологически. Еще одна особенность этого локуса была обнаружена при скрещиваниях между самими линиями, сбалансированными по летал ям. Обычно скрещивание гетерозиготы T/t a с такой же гетерозиготой дает только бесхвостых потомков. Однако изредка (1:500 - 1:1000) среди потомков встречаются мыши с нормальными хвостами. Эти нормальные мыши почти всегда получаются в результате редкой генетической рекомбинации в 17-й хромосоме, в участке, соответствующем локусу Т или примыкающем к нему. Можно показать, что наличие хвоста у этих реком-бинантных потомков обусловлено тем, что одновременно с рекомбинацией произошло превращение первоначального аллеля t a в новый аллель t x , который комплементарен аллелю t a таким же образом, как некоторые t -аллели, происходящие от мышей дикого типа, комплементарны друг друга. Различие, конечно, состоит в том, что t x в данном случае непосредственно связан с t a своим происхождением. Было показано, что такое превращение одного аллеля t в другие комплементарные типы происходит с большинством выделенных рецессивов. Некоторые возникающие при этом аллели, например t x , могут в свою очередь при помощи того же механизма превратиться в другой комплементарный аллель, например t y. Такое превращение одного рецессивного аллеля t в другой приводит к образованию ряда постепенно переходящих один в другой аллелей, а в конечном итоге к созданию так называемых t viable, или t v , аллелей. Все эти последние аллели нелетальны, и бесхвостый фенотип экспрессируется только в случае гетерозигот Т/t v; у особей t v/t v хвосты нормальные. Еще один класс рецессивных t -мутаций - это полулетали, жизнеспособность которых колеблется от 2 до 51% нормальной. Как и полностью летальные аллели, они путем постепенных превращений дают t v -типы. Комплементарные скрещивания ( T/t a x T/t b) всех как природных, так и полученных экспериментально рецессивных аллелей показали, что 111 существующих мутаций распадаются на 8 отдельных групп, причем ни одна из них не комплементарна Т -аллелю. Число членов во всех группах различно - от одного в группе ( t w73до 66 в группе аллелей t v . Из пяти доминантных Т -мутаций одна была вызвана воздействием рентгеновских лучей, а все другие мутации, обнаруженные в этом локусе, по-видимому, возникли спонтанно.
Мы рассмотрели несколько особенностей этого сложного ряда генетических дефектов, связанных между собой либо тем, что они комплементарны и определяются соседними локусами, либо тем, что они происходят друг от друга. Перейдем теперь к эмбриологическим особенностям экспрессии сложного локуса.
Каждая из восьми комплементарных групп обусловливает разного рода дефекты, проявляющиеся в период от ранних до поздних стадий развития зародыша. Морфологические проявления этих дефектов представлены на рис. 7-10 и 7-11. Самые ранние дефекты обнаружены у зародышей, гомозиготных по аллелю t 12. Оплодотворение и деления дробления зиготы приводят к образованию шаровидной массы клеток, называемой морулой. Первый признак клеточной дифференцировки у мышей наблюдается при переходе от этой морулы к следующей стадии - бластоцисте, состоящей из трофобласта (трофэктодермы) и внутренней клеточной массы. Гомозиготные t 12-зародыши не достигают этой стадии и недифференцированные «морулы» гибнут, не имплантируясь в стенку матки, в отличие от нормальных зародышей, которые делают это примерно на 4-й день после оплодотворения. Кроме того, t 12-клетки, по-видимому, автономны в своем летальном действии. Химеры, содержащие эмбриональные клетки t 12и нормальные эмбриональные клетки, летальны, и их развитие не заходит дальше той стадии, на которой гибнут зародыши t 12. Поэтому мутация t 12, по-видимому, затрагивает локус, необходимый для первого шага в дифференцировке мышиного зародыша - возникновения трофобласта, из которого в конечном итоге формируются хорион и другие характерные для всех плацентарных млекопитающих внезародышевые оболочки, образующиеся из зиготы.
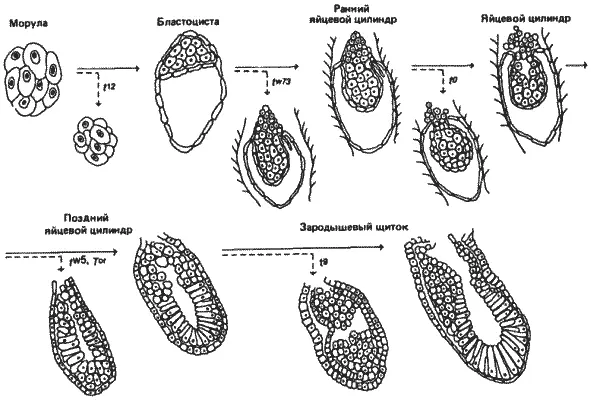
Рис. 7-10.Схематическое изображение событий, происходящих на ранних стадиях развития зародыша мыши. Сплошными стрелками показано течение нормального онтогенеза, а прерывистыми - момент отклонения от нормального развития зародышей, несущих различные t -аллели. Более подробные объяснения см. в тексте (Bennett, 1975; с изменениями).
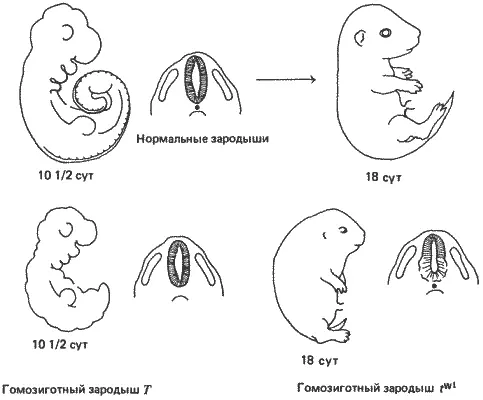
Рис. 7-11.Мышиные зародыши на поздних стадиях развития, иллюстрирующие эффекты двух t -аллелей, действующих на поздних стадиях. Подле каждого зародыша приведена схема поперечного сечения, на которой показаны нервная трубка (заштрихована), хорда (черная точка под нервной трубкой) и сомиты (продолговатые структуры по обе стороны нервной трубки). Полное описание мутантных фенотипов дано в тексте (Bennett, 1975).
Читать дальшеИнтервал:
Закладка:









