Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Следующий из аллелей, вступающих в действие на очень ранних стадиях, - это аллель t w73. Зародыши, гомозиготные по этому аллелю, образуют бластоцисту. Однако трофобласт таких мутантных зародышей не вступает в надлежащую связь со стенкой матки, и недостаточно хорошо имплантировавшийся зародыш вскоре гибнет.
В норме после успешной имплантации зародыша в стенку матки внутренняя клеточная масса начинает расти и подвергается дальнейшей дифференцировке. Одно из происходящих при этом событий - формирование внезародышевой и зародышевой эктодермы. Из первой в конечном счете образуются плацента и части внезародышевых оболочек, а из второй - собственно зародыш. Мутанты t 0не образуют внезародышевую эктодерму и гибнут на стадии раннего яйцевого цилиндра.
Далее у нормальных зародышей продолжается рост внутренней клеточной массы и образуется продолговатое скопление эктодермальных клеток, покрытое энтодермой и называемое яйцевым цилиндром. Внезародышевые клетки также продолжают пролиферировать и дифференцироваться. Зародыши, гомозиготные по аллелю t w5, доходят до стадии, похожей на яйцевой цилиндр, после чего клетки зародышевой эктодермы подвергаются пикнозу и гибнут. Клетки внезародышевой эктодермы, по-видимому, остаются незатронутыми и на протяжении нескольких дней продолжают нормально развиваться, несмотря на находящийся внутри мертвый зародыш, но в конце концов также гибнут.
На этой стадии развития мышиного зародыша, достигаемой через 6,5-7 сут после оплодотворения, начинается дифференцировка собственно зародыша. Это проявляется в формировании на яйцевом цилиндре первичной полоски и образовании между уже имеющимися зародышевыми эктодермой и энтодермой слоя мезодермальных клеток. У зародышей, гомозиготных по аллелю t 9, нормальная мезодерма не образуется и соответственно не образуется ни одно из ее производных. Поскольку мезодермальные клетки образуются из зародышевой эктодермы в области первичной полоски, можно предположить, что мутантные эктодермальные клетки неспособны к такому превращению. Мутантные t 9-зародыши, пересаженные в брюшную полость нормальных взрослых особей, образуют злокачественные опухоли, целиком состоящие из эктодермальных тканей.
Первичная полоска - это место образования трех зародышевых листков и развития первичной оси зародыша.
По обе стороны первичной оси на большей части ее длины образуются сомиты, а медиально по отношению к парным сомитам - хордомезодерма. Последняя индуцирует лежащую над ней нейральную эктодерму к формированию нервной трубки - структуры, которая в конечном итоге становится спинным мозгом, а на переднем конце головным мозгом. Все эти события происходят и у зародышей t w 1. Однако после образования нервных структур вентральная часть нервной трубки и головной мозг у них дегенерируют. Сохраняющиеся дорсальные клетки физически замещают мертвые вентральные клетки, но, по-видимому, не могут заменить их функционально, потому что у этих мутантных зародышей всегда наблюдаются разного рода дефекты и они гибнут до рождения.
Конечное летальное состояние, обусловленное локусом Т , можно видеть у особей, гомозиготных по доминантному аллелю Т . Летальная фаза у этих особей сходна с наблюдаемой у мутантов t w 1, т. е. она наступает позднее, чем в случае большинства других рецессивных аллелей. У особей Т/Т первичная полоска не достигает заднего конца зародыша. Поэтому ни одна из структур, зависящих от формирования мезодермы, в этой области никогда не развивается. Кроме того, что более важно, аллантоисная ножка плаценты у них не образуется, т.е. зародыш лишается необходимых для его жизни нормальных связей с плацентой. Вдобавок не развивается ни одна из структур самого зародыша, находящихся позади почек передних конечностей. Наконец, несмотря на, казалось бы, нормальный передний конец первичной полоски и на образование сомитов и хорды, эти последние структуры не сохраняются, а исчезают. В результате строение переднего конца зародыша оказывается сильно нарушенным.
Создается впечатление, что, хотя дефекты, наблюдаемые у этой галереи уродцев, развивающихся под действием мутаций в сложном локусе Т , очень разнообразны, все они имеют одну общую особенность. Как отметил Беннетт (Bennett), все летальные Т-аллели вызывают дефекты эктодермы. Эти дефекты выражаются в нарушении способности эктодермы мутантов либо нормально дифференцироваться ( t 12и t 0), либо нормально функционировать ( t w73и t w1). Описанные выше дефекты схематически представлены на рис. 7-12. Как показано на схеме, можно представить себе, что различные мутации локуса Т должны делать ряд последовательных выборов из двух возможностей, определяющих дальнейшую судьбу эктодермы и всех ее производных. Первоначально морула состоит из недифференцированных клеток. Мутация t 12препятствует принятию первого решения - выбору между трофобластом и внутренней клеточной массой. Затем мутация t w73нарушает надлежащее функционирование трофобласта. Мутация t 0препятствует образованию внезародышевой эктодермы; ее эффект, возможно, аналогичен эффекту мутации t 12. На параллельном этапе мутация t w5убивает эктодерму собственно зародыша. Наконец, мутации t 9, t w1и Т нарушают различным образом либо способность зародышевой эктодермы дифференцироваться в мезодерму, либо способность мезодермы, если она уже образовалась, индуцировать или поддерживать нервную ткань.
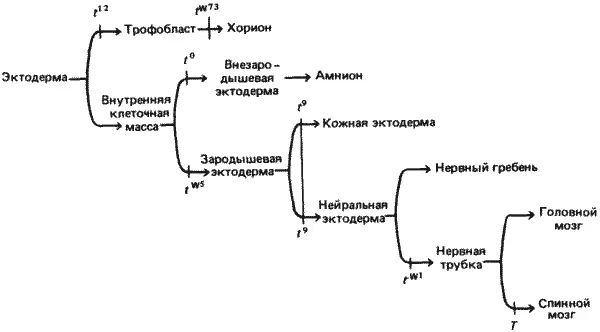
Рис. 7-12. Предполагаемая цепь последовательных решений, которые должны приниматься в процессе дифференцировки эктодермы и ее производных у мыши. Над стрелками указаны различные t -аллели, блокирующие соответствующие процессы.
Первичное нарушение, лежащее в основе всех этих далеко идущих эффектов всего лишь одного локуса, составляет в настоящее время предмет многочисленных споров и экспериментальных исследований. Однако, какова бы ни была непосредственная причина (или причины) этих дефектов, совершенно очевидно, что локус Т играет первостепенную роль в морфогенезе одного из трех зародышевых листков мышиного зародыша. Поэтому он, подобно локусу Notch у дрозофилы, принимает важное участие в развитии организма в целом.
Мутации, воздействующие на развитие определенных органов
В то время как локус Т оказывает, по-видимому, самые разнообразные воздействия на все развитие эктодермы, существуют мутации, вызывающие более специфические дефекты. Примером служит мутация cardiac lethal (с) у аксолотля Ambystoma mexicanum. Эта мутация была впервые обнаружена и исследована Хэмфри (Humphrey). Хэмфри установил, что мутация с наследуется как простой аутосомный рецессивный признак, так что при скрещивании двух гетерозиготных особей (с/+) 25% потомков гибнет на ранних личиночных стадиях, вскоре после вылупления. Эти мутантные особи плавают, как нормальные личинки, но они раздуты переполняющей их жидкостью, а пищеварительная система и жабры у них недоразвиты (рис. 7-13). Первичная причина этих дефектов - нарушение развития сердца и его неспособность к сокращениям. Поэтому у мутантных личинок отсутствует кровообращение, а дышат они, вероятно, путем диффузии через кожу, что дает им возможность просуществовать лишь в течение ограниченного периода времени. Как показал Хэмфри путем сращивания нормального и мутантного зародышей, это нарушение развития сердца носит автономный характер.
Читать дальшеИнтервал:
Закладка:









