Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Эксперимент Хэмфри схематически представлен на рис. 7-14. У мутантного и нормального зародышей, взятых до закладки сердца, удаляли по кусочку ткани с боковых поверхностей тела. Затем зародышей соединяли по месту раны и давали им срастись. В тех случаях, когда такие сросшиеся особи завершали развитие, было установлено, что нормальный партнер ослаблял раздутость и другие дефекты зародыша с/с, давая ему возможность выжить. Однако сердце мутантного партнера навсегда оставалось простой трубкой, неспособной к сокращениям, и кровообращение мутанта целиком обеспечивал нормальный партнер.
Установлено, что у позвоночных многие органы, в том числе сердце, развиваются в результате определенных индукционных взаимодействий, происходящих во время развития. В частности, Джекобсон и Дункан (Jacobson, Duncan) показали, что у хвостатых амфибий развитие сердца из мезодермальных зачатков индуцируется головным участком энтодермы. Причиной того, что особям с/с не удается образовать сердце, может быть неспособность головного участка энтодермы индуцировать этот процесс или же неспособность сердечной мезодермы реагировать на индуктор. Для того чтобы выяснить, какая из этих двух причин вызывает нарушение развития,
Хэмфри пересаживал нормальную сердечную мезодерму с/с -реципиентам, а мезодерму с/с -мутантов - нормальным реципиентам. Оказалось, что мезодерма с/с способна образовать сокращающееся сердце под индукционным воздействием нормального головного участка энтодермы, тогда как мутантные зародыши не могут обеспечить развитие нормального сердца. Эти результаты можно интерпретировать как указание на отсутствие индукционной активности головного участка энтодермы у мутанта с/с . Однако возможно также, что мутантные особи активно подавляют формирование сердца. Эксперименты, проведенные Лемански (Lemanski) и его сотрудниками, делают последнее предположение менее вероятным. Эти авторы выращивали in vitro мезодермальные зачатки сердца мутантных и нормальных зародышей. В использованных ими условиях в нормальной сердечной мезодерме происходили сильные сокращения, а в мутантной их не было. Если считать, что сокращения мутантной ткани подавлялись in situ , то выращивание in vitro должно было снять это воздействие. Более того, при совместном культивировании мезодермы мутанта с/с и головного участка энтодермы нормального зародыша в мутантной ткани начинались сокращения; это показывает, что мутантная мезодерма способна нормально реагировать на соответствующее индукционное воздействие. Таким образом, ген «cardiac lethal», очевидно, обусловливает неспособность головного участка энтодермы обеспечить индукционный сигнал, запускающий дифференцировку сердца из его мезодермального зачатка.

Рис. 7-13.Нормальная личинка аксолотля Ambystoma mexicanum (А) и мутантная личинка (Б), гомозиготная по аллелю с. Тело мутантной личинки раздуто вследствие накопления избыточной жидкости (Kulikovski, Manasek, 1978).
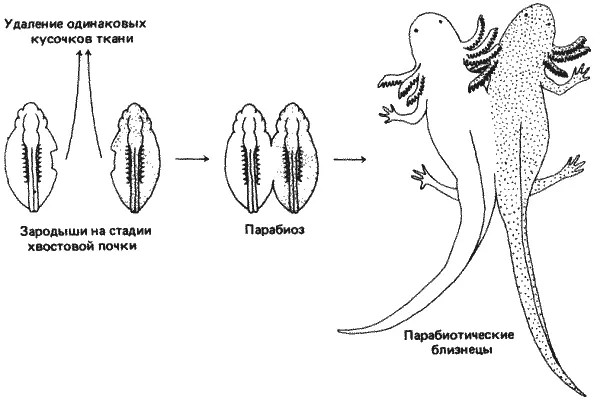
Рис. 7-14.Схема метода парабиоза. Двух аксолотлей, различающихся по генотипу (показано наличием или отсутствием точек), сращивают на ранних стадиях эмбриогенеза. Как показывает дальнейшее развитие, нормальная особь способна «спасти» своего мутантного партнера.
Исследования, проведенные на другой аутосомной рецессивной мутации аксолотля - eyeless (e), дали результаты, прямо противоположные полученным на cardiac lethal. Как показывает ее название, особи, гомозиготные по этой мутации (е/е), лишены глаз. Она вызывает также два других плейотропных дефекта - темную пигментацию и стерильность. Основной дефект возникает на самых ранних стадиях развития глаз, которое блокируется в начале формирования глазных пузырей или до этого. Подобно сердцу, глаз формируется в результате индуктивного взаимодействия, происходящего между передним участком нейральной эктодермы в презумптивном переднем мозге (передняя медуллярная пластинка) и хордомезодермой, располагающейся под этим участком во время гаструляции. Индуцированная нервная ткань образует боковые выросты, которые в конце концов инвагинируют, образуя глазные бокалы. Ван Дейзен (Van Deusen) исследовал природу дефекта, обусловливаемого мутацией е, пересаживая прехордальную мезодерму из спинной губы бластопора мутантных и нормальных зародышей в бластулы. При пересадке мезодермы е/е в нормальные бластулы она индуцировала развитие глазных пузырей, пересадка же нормальной мезодермы в бластулы е/е не приводила к индукции раннего развития глаза. Такие же результаты были получены при реципрокных трансплантациях, если кусочки мезодермы брали в более поздние сроки ранней гаструляции, но до формирования глазных пузырей: нормальная эктодерма была способна образовать глазные пузыри под действием как нормальной, так и е/е -мезодермы, а эктодерма е/е не индуцировалась ни той, ни другой мезодермой. Поэтому можно полагать, что дефект eyeless вызывается, очевидно, неспособностью эктодермы реагировать на мезодермальный индуктор. В пользу такого заключения свидетельствует также то, что если пересадить морфологически дифференцированный глаз от нормальной особи к е/е -особи, то он приживается и функционирует. Таким образом, после того как глазной пузырь индуцирован, генотип е/е способен поддерживать полностью сформированный глаз. Однако два других плейотропных дефекта мутантов е/е - пигментация и стерильность - все еще требуют объяснения. Избыточно пигментированный фенотип можно без труда создать экспериментально, удалив у нормальной особи глазные пузыри; это приводит к развитию у слепой личинки очень сильной пигментации. И наоборот, если пересадить развивающейся особи е/е зачаток нормального глаза, то ее пигментация становится нормальной. Это показано на рис. 7-15, на котором представлены фотографии нормальной личинки, мутанта е/е и одноглазой личинки е/е. Избыточная пигментация - следствие одного лишь наличия или отсутствия глаз, а поэтому она обусловлена относительной плейотропией.
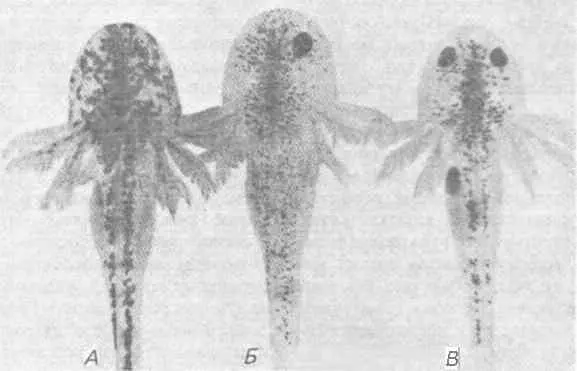
Рис. 7-15.Личинки аксолотля: мутант eyeless (А), особь с генотипом eyeless, которой был пересажен глаз от нормальной особи ( Б ), и нормальная особь ( В ). Пересадка привела к ослаблению пигментации по сравнению с мутантной особью (А) и приблизила ее к нормальному фенотипу ( В ) (Ерр, 1978).
Читать дальшеИнтервал:
Закладка:









