Ирина Спивак - Репликация ДНК: учебное пособие

- Название:Репликация ДНК: учебное пособие
- Автор:
- Жанр:
- Издательство:Array Издательство Н-Л
- Год:2011
- Город:СПб.
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Ирина Спивак - Репликация ДНК: учебное пособие краткое содержание
В пособии описываются проблемы репликации ДНК. Излагаются современные представления о строении хромосом, координации в течение клеточного цикла процессов ДНК-метаболизма, а также описываются участвующие в этих процессах белки и рассматриваются механизмы, отвечающие за сохранение генетической стабильности организмов.
Предназначено для студентов дневной, очно-заочной и заочной форм обучения, изучающих дисциплины «Экология» и «Физико-химические основы цитологии» в рамках подготовки бакалавров по направлению 140400 «Техническая физика».
Репликация ДНК: учебное пособие - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
2.2. Ориждин репликации E.coIi oriC
Наиболее подробно изучены ориджины у Е. соli и Bacillus subtilis. Область начала репликации хромосомы, оriС (origin of chromosome), включает в себя участки со специфическими последовательностями, так называемыми ДНК-боксами, и расположенными между ними короткими последовательностями. ДНК-боксы со специфическим «мотивом» нуклеотидов, преимущественно в 9пн, перемежаются фрагментами в 12-1Зпн с высоким содержанием АТ. Сами девятичленные последовательности могут располагаться как в прямом, так и в инвертированном положении по отношению друг к другу. Например, у В. subtilis имеется один фрагмент ТТАТССАСА и два других девятичленных бокса, ориентированных в противоположном направлении, с заменой одной из пар нуклеотидов. Всего у В. subtilis на оriС расположено 15 ДНК-боксов. Область оriС очень консервативна: ДНК-боксы сходного состава имеются в соответствующем месте хромосомы у других бактерий (только у Mycoplаsma genitalium, несмотря на наличие общих для всех бактерий ферментов репликации, ДНК-боксов найдено не было). Сами ДНК-боксы не кодируют белок или РНК, хотя между ними располагаются отдельные гены. Продукты этих генов также большей частью вовлечены в «обслуживание» процесса репликации ДНК.
Порядок расположения ДНК-боксов, промежуточных областей и их количество позволяют думать, что эволюционная дивергенция oriС шла главным образом за счет дупликаций и трипликаций. Схема абстрактного «минимального ориджина» прокариот представлена на рис. 3.
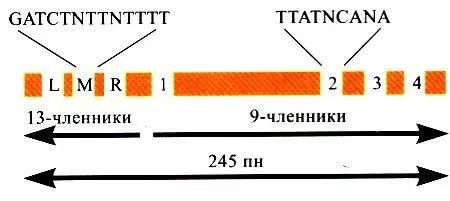
Рис. 3.Организация минимального ориджина прокариот
Схема минимального ориджина прокариот.
2.3. Ориджины других организмов
Коровая часть ориджина репликации у вируса SV40 состоит из элемента опознания (ORE – origin recognition element), необходимого для связывания особого белка Т-антигена (Т-аg), элемента для связывания белка, расплетающего ДНК (DUE – DNA unwinding element), и элемента, обогащенного АТ-нуклеотидами. Участок, с которого вилка репликации начинает двигаться в противоположных направлениях, называется началом двунаправленной репликации (OBR – origin bidirectional replication).
Вспомогательные элементы (Aux) связывают димеры Т-антигена (Аux-1) и фактор транскрипции Sp1 (Аuх-2). Расстояние между этими элементами и их ориентация играют важную роль в процессе инициации репликации. Схема ориджина вируса SV40 представлена на рис. 4.
У эукариот гомологами ориджинов репликации являются автономно реплицирующиеся последовательности, или ARS (autonomously replicating sequences), открытые в 1980 г. Р. Дэйвисом и Дж. Карбоном.
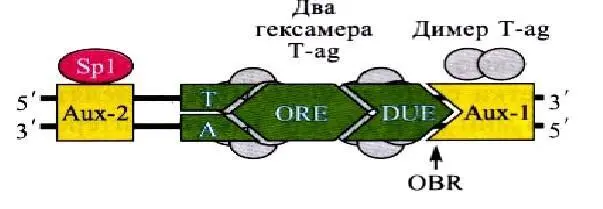
Рис. 4. Схема ориджина вируса SV40.
У дрожжей Saccharomyces cerevisiae особые последовательности, способные обеспечивать репликацию фрагментов ДНК в дрожжевой клетке были выделены раньше, чем у других эукариот. Позднее такие последовательности были найдены и у многих других организмов. У S.cerevisiae АRS занимает 100—200пн и содержит специфическую консенсусную последовательность (АСS – ARS consensus sequence), размером в 11пн, необходимую для связывания с белком-инициатором, а также дополнительные элементы (В-элементы), усиливающие функцию ориджина. Например, АRS1 – первый подробно охарактеризованный ориджин – содержит три таких элемента – В1, В2, ВЗ. Последовательности АCS и В1 занимают приблизительно 50пн и представляют собой наименьшую функциональную область любого ориджина, которая требуется для связывания с белком-инициатором.
Элемент В2 обычно содержит генетически охарактеризованный участок DUE. Вспомогательный элемент ВЗ связывает фактор транскрипции Abf-1. Общая длина ARS-элемента составляет 100-200пн. Строение ориждина S.cerevisiae представлено на рис. 5.
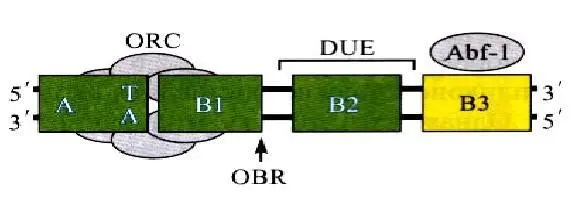
Рис. 5. Схема ориджина Saccharomyces cereiseae
У другого вида дрожжей, Shizosaccharomyces pombe, ориджины состоят по крайней мере из одной ARS, которая значительно длиннее, чем у S. cerevisiae. В некоторых случаях несколько ARS-элементов формируют зону инициации репликации. (Рис. 6.)

Рис. 6. Схема ориджина Shizosaccharomyces pombe
У млекопитающих ориджины детально не охарактеризованы, некоторые из них располагаются в межгенных промежутках, имеют сайты связывания для транскрипционных факторов, часто содержат только районы инициации двунаправленной репликации – OBR.
2.4. Скорость репликации
Скорость репликации генома регулируется в основном частотой инициирующих событий. Так, у Е. соli скорость копирования в каждой репликативной вилке постоянна и равна примерно 1500пн в секунду: следовательно, полный геном длиной 4·10 6пн реплицируется примерно за 40 мин. Если хромосома реплицируется быстрее, это значит, что увеличивается частота актов инициации в той же самой точке начала репликации при прежней скорости копирования. Клетки Е. соli делятся каждые 20 мин; это означает, что репликация ДНК инициируется в хромосомах, еще не закончивших предыдущий раунд репликации. Скорость движения репликативной вилки в эукариотических клетках значительно меньше (10-100пн в секунду), но завершение репликации хромосомы в разумное время обеспечивается одновременной инициацией во множестве точек. Итак, скорость репликации хромосом контролируется числом и расположением точек начала репликации. Например, в ранних эмбрионах дрозофилы репликация отдельной хромосомы осуществляется каждые 3 мин, благодаря почти одновременной инициации событий в точках, отстоящих друг от друга на 7000-8000пн. В тоже время известно, что у дрозофилы в ходе раннего эмбрионального развития, как скорость репликации, так и размеры и число репликонов тканеспецифичны. В культуре же соматических клеток той же дрозофилы скорость удвоения хромосом значительно более медленная, так как репликация начинается в гораздо меньшем числе точек, находящихся друг от друга на расстоянии 40000пн, при этом продолжительность S-фазы составляет 600 мин. Следовательно, при фиксированной скорости синтеза ДНК множественная инициация повышает скорость процесса репликации в целом и таким образом уменьшает время, необходимое для удвоения всего набора хромосом. Данные о числе репликонов и скорости репликации приведены в табл.1.
Различия в продолжительности S-фазы найдены и у других организмов. Например, у тритона S-фаза длится 1 ч в ядрах бластулы и 200 ч в предмейотической S-фазе сперматоцитов. Вероятно, длительность S-фазы определяется не скоростью синтеза ДНК, а числом задействованных ориджинов репликации. В ДНК клеток нейрулы тритона они находятся на расстоянии около 40 мкм друг от друга, а в соматических клетках – около 100 мкм.
Читать дальшеИнтервал:
Закладка:









