Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Жанр:
- Издательство:Литагент БИБКОМ
- Год:2012
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных краткое содержание
Общая вирусология с основами таксономии вирусов позвоночных - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Мембраны клетки-хозяина. В отличие от фаговых репликаз, RdRp вирусов эукариот неизменно связана с надмолекулярными структурами: мембранами клеткихозяина у (+)РНК-вирусов, нуклеокапсидом у (-)РНК-вирусов и субвирусными частицами у днРНК-вирусов. Внутриклеточные мембраны клеток, инфицированных вирусами с (+)РНК-геномом, подвергаются быстрому перераспределению, формируя места заякоривания вирусных репликативных комплексов. Когда эти комплексы отсоединяются от мембран, они теряют способность катализировать истинную репликацию РНК, хотя часто сохраняют ограниченную способность копировать РНК-матрицу. При изучении нодавирусной инфекции истинная РНК-репликазная активность частично очищенной RdRp была восстановлена путем добавления к бесклеточному экстракту глицеролфосфолипидов. Эти результаты подтвердили идею, что мембранная организация играет центральную роль в репликации (+)РНК. То же самое заключение получено при ингибировании репликации РНК полиовируса брефелдином А, который блокирует внутриклеточные мембранные взаимодействия. Хотя определенная роль мембран неясна, вероятно, они могут ускорять сборку репликативных комплексов, сокращая время процесса и отделяя дочерние молекулы от матриц.
Механизмы репликации РНК-геномов. Как уже отмечалось, репликация РНК-геномов осуществляется вирусоспецифической RdRp, которая может входить в состав вириона или детерминироваться геномом. В отличие от ферментов, которые копируют ДНК с использованием затравки, большинство RdRp могут начать синтез РНК de novo . Исключением является RdRp пикорнавирусов, которая для инициирования синтеза использует маленький вирусный белок (VPg), ковалентно связанный с урацилом. VPg удаляется при трансляции генома, но сохраняется при его инкапсидации.
Интересно, что у тогавирусов (вирус Синдбис), репликация (+)РНК на стадии синтеза минус-нити (образование РФ) осуществляется только переходной версией RdRp, которая впоследствии протеолитически процессируется, что переключает матричную специфику RdRp на синтез положительных нитей.
Репликациция (+)РНК полиовирусов .
Полиовирусы – мелкие (27 нм) безоболочечные икосаэдрические вирусы, поражающие позвоночных. Геном – линейная однонитевая РНК позитивной полярности. На 5'-конце РНК ковалентно связана с терминальным геномным белком через остаток тирозина, 3'-конец полиаденилирован (рисунок 8).
Репликацию/транскрипцию генома осуществляет РНК-полимераза, детерминированная 3'-концом генома, который транслируется сразу после попадания вируса в клетку. На первой стадии репликации происходит образование двухнитевой РФ за счет синтеза минус-нити, инициированного присоединением молекулы урацила к 3'-поли-А концу.
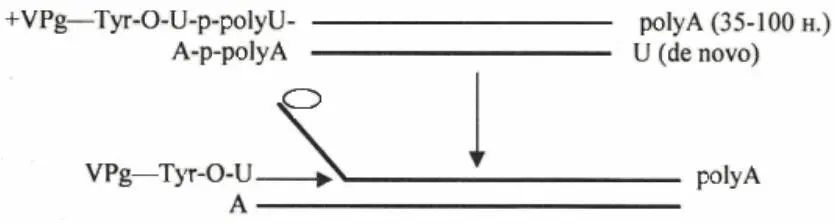
Рисунок 8 – Схема репликации РНК полиовирусов
Кроме РНК-полимеразы в клетке синтезируется вирус-специфический терминальный низкомолекулярный белок (VPg), который через тирозин связывается с молекулой урацила. Данная структура используется РНК-полимеразой в качестве затравки – то есть происходит терминальная инициация с использованием нуклеотидбелковой затравки. Синтез идет с вытеснением цепи. Образующиеся молекулы (+)РНК до накопления достаточного количества вирусоспецифических белков используются как мРНК, после чего они начинают инкапсидироваться в вирусную частицу.
Следует отметить, что представленная схема репликации геномной (+)РНК полиовирусов не является универсальной. Вирусные (+)РНК-геномы различаются организацией 5'– и 3'-концевых структур, что определяет особенности их репликации, связанные с инициацией синтеза.
Репликациция (-)РНК-геномов .
Вирусные (-)РНК-геномы могут быть непрерывными или сегментированными. Во всех случаях РНК находится в составе рибонуклеопротеина, что и определяет особенности ее репликации, поскольку депротеинизированная РНК не может служить матрицей для полимеразы. Все вирусы с (-)РНК-геномом имеют собственную РНК-зависимую РНК-полимеразу, входящую в состав РНП. Для получения полноразмерного генома должна быть синтезирована репликативная полноразмерная плюс-нить. Однако на первом этапе репродуктивного цикла геномная (-)РНК служит матрицей для транскрипции, которая протекает с последующим процессингом мРНК, и не может служить матрицей для синтеза полноразмерной копии. Синтез репликативной полноразмерной (+)РНК начинается только после накопления соответствующих вирусных белков, подавляющих преждевременную терминацию РНК на внутренних участках матрицы. Каким образом это происходит, остается пока неизвестным. Синтез антигеномной и геномной РНК происходит в составе РНП.
Репликациция днРНК реовирусов .
Реовирусы – двукапсидные (60-75 нм) частицы с икосаэдрическим типом симметрии, инфицируют позвоночных, беспозвоночных, растения. Геном состоит из 10-12 фрагментов днРНК.
Репликация днРНК неразрывно связана с транскрипцией, которая является ее первой стадией.
1 Синтез (+)РНК на двухнитевой матрице протекает по консервативному типу без вытеснения цепи и происходит в составе однокапсидной вирусной частицы при участии белков кора – вирусной РНК-зависимой РНК-полимеразы (VP2) и гуанидилтрансферазы (VP3). мРНК покидают частицу через поры внутреннего капсида.
2 Плюс-нити РНК объединяются с вновь синтезированными белками кора и неструктурными (NS) белками. При созревании вириона РНК-полимераза осуществляет синтез минус-нитей на матрице (+)РНК по репарационному механизму, затягивая ее внутрь формирующегося капсида. Сформированная однокапсидная частица может снова начать синтез плюс-нитей РНК.
Как и в случае полиовирусов, представленный способ репликации генома реовирусов не является универсальным для вирусов с днРНК геномом. Например, у фага φб синтез плюс-нитей на родительском дуплексе происходит по полуконсервативной модели и всегда сопряжен с вытеснением предшествующей нити (+)РНК.
3.7.1.3 Основные принципы и механизмы репликации ДНК-геномов вирусов
В процессе репликации ДНК-содержащие вирусы осуществляют некоторые шаги, которые отсутствуют у РНК-геномных вирусов. Для большинства ДНК-содержащих вирусов генетические стратегии включают: транспорт ДНК вириона в ядро клетки, инициирование транскрипции с этой ДНК, индукцию транскрипции дополнительных вирусных генов, подготовку клетки для репликации ДНК вируса, дублирование ДНК-генома, упаковку ДНК в вирионы и выход вирусных частиц из ядра. Кроме этого, многие ДНК-вирусы развили уникальные механизмы уклонения от иммунной защиты организма и способность вызывать опухоли у животных. В процессе близких отношений со своими хозяевами, вирусы эксплуатируют ключевые клеточные регуляторные системы и узурпируют важные клеточные процессы. В связи с этим, изучение различных аспектов репликации ДНК-вирусов обеспечивает новые фундаментальные знания о молекулярных процессах, происходящих в клетке, включая выражение генов, репликацию ДНК и контроль за циклом клеточного деления.
Читать дальшеИнтервал:
Закладка:









