Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Жанр:
- Издательство:Литагент БИБКОМ
- Год:2012
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных краткое содержание
Общая вирусология с основами таксономии вирусов позвоночных - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
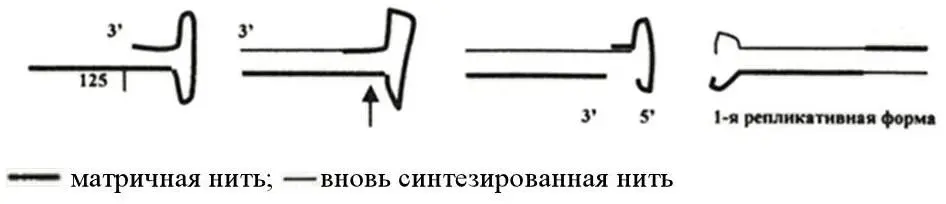
Рисунок 10 – Схема первых этапов репликации однонитевой ДНК парвовирусов
Концевые 125 нуклеотидов родительского генома становятся условной частью вновь синтезированной цепи и возникший таким образом 3’-конец родительской цепи используется для ее регенерации. В результате этих реакций возникает дисперсная двунитевая репликативная форма вирусной ДНК (рисунок 10). Далее следует цепь реакций, включающих образование на одном из концов ДНК-затравки в виде «заячьих ушек», синтез новой цепи с вытеснением родительской, образование еще одной репликативной формы. Вторая репликативная форма ДНК используется в качестве матрицы для дальнейшего синтеза вирусной ДНК, а вытесненная из дуплекса однонитевая молекула или вступает в репликативный цикл или входит в состав дочерней вирусной частицы.
2 Репликация с использованием терминальной инициации при помощи белокнуклеотидной затравки (рисунок 11). Такой тип репликации геномной ДНК имеют аденовирусы, геном которых представлен линейной днДНК, имеющей на 5’-концах инвертированные повторы и ковалентно присоединенные геномные белки с м.м. 55 кДа.
В инфицированной аденовирусом клетке синтезируется вирусоспецифический белок массой 80 кДа, который связывается через серин с дезоксицитидином. Образовавшаяся структура Б-Ser – dCTPявляется затравкой, которая через цитозин комплементарно связывается с 3’-концевым гуанозином генома и инициирует синтез цепи ДНК.
Инициация может наблюдаться на любом конце родительской ДНК и может происходить или одновременно или последовательно. При последовательной инициации синтез дочерней цепи сопровождается вытеснением одной из родительских, а синтез комплементарной цепи идет на однонитевой матрице по репарационному механизму. В тоже время обсуждается и другой механизм синтеза второй нити. Замещенная родительская однонитевая ДНК имеет на концах самокомплементарные инвертированные повторы, которые отжигаются, восстанавливая двунитевую точку ori, узнаваемую инициирующими белками, обеспечивающими синтез родительскодочернего дуплекса. Таким образом, каждый родительский дуплекс копируется полуконсервативно.
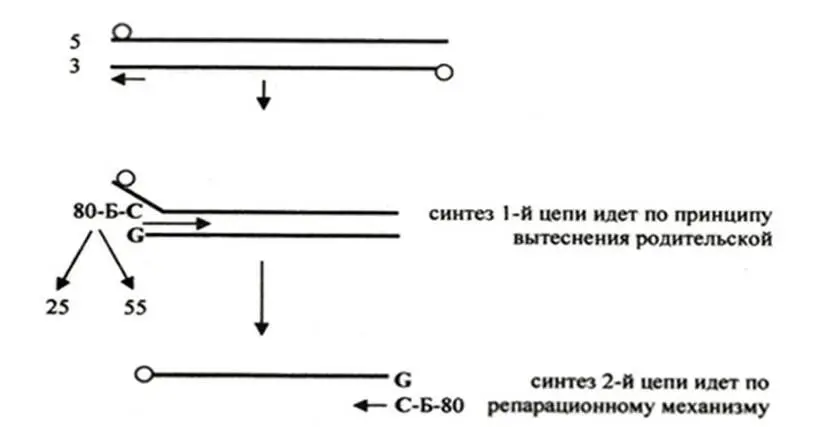
Рисунок 11 – Схема репликации генома аденовируса
Однако процесс протекает без синтеза отстающей цепи, т.е. без образования множественных сайтов инициации и синтеза фрагментов Оказаки.
3 Репликация кольцевых геномов по механизму катящегося кольца (рисунок 12). Катящееся кольцо – способ репликации, при котором репликационная вилка совершает множество оборотов на кольцевой матрице. Синтезирующаяся в каждом цикле нить вытесняет прежнюю (гомологичную) цепь двуцепочечной молекулы, синтезированную в предыдущем цикле, образуя хвост, состоящий из набора последовательностей, комплементарных одноцепочечному матричному кольцу. В общих чертах репликация по механизму катящегося кольца имеет следующие стадии:
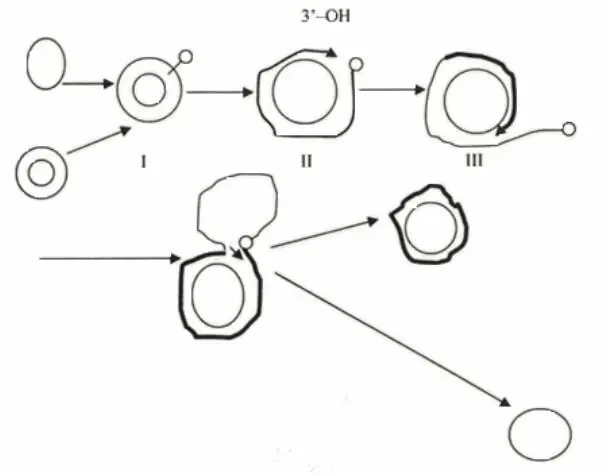
Рисунок 12 – Схема репликации ДНК-геномов по механизму катящегося кольца
1 Вирусоспецифический фермент вносит однонитевой разрыв в уникальном сайте родительской цепи репликативной формы.
2 Фермент остается связанным с 5’-концом, освободившийся 3’-концевой нуклеотид служит затравкой для ДНК-полимеразы.
3 ДНК-полимераза присоединяет нуклеотиды комплементарно замкнутой цепи, то есть синтезируется только лидирующая цепь. 5’-конец родительской цепи вытесняется. Наблюдается образование сигма-молекул (δ).
4 После того, как репликационная вилка завершит чуть больше полного оборота, вытесненная цепь замыкается в кольцо, а фермент перемещается на вновь синтезированную нить и цикл повторяется. Таким образом, вновь синтезированная нить, имеющая последовательность геномной, становится компонентом РФ, а предшествующая (родительская) оказывается в свободном виде.
Механизм катящегося кольца при репликации ДНК используют многие бактериофаги.
Однако этот механизм не игнорируют и вирусы эукариот. Например, линейная вирионная ДНК вируса герпеса при попадании в клетку переходит в кольцевую форму и, пройдя первую стадию тета-репликации (см. ниже), реализует механизм катящегося кольца.
Однако, вместо производства кольцевых дочерних молекул, репликация генерирует конкатамерные молекулы. Чтобы восстановить линейные дочерние молекулы ДНК вирусоспецифические белки расщепляют конкатамеры в определенных сайтах последовательности в процессе упаковки ДНК в капсиды.
4 Репликация ДНК по схеме Кернса (тета-репликация) включает несколько этапов (рисунок 13):
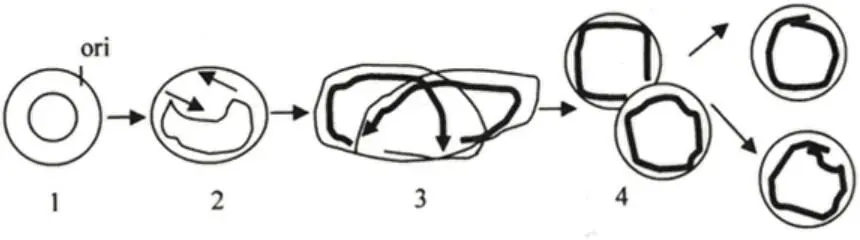
Рисунок 13 – Репликация ДНК по схеме Кернса
1) вирусоспецифический неструктурный белок, обладающий хеликазной активностью, связывается с ДНК-последовательностью в точке ori и расплетает двунитевую структуру;
2) праймаза синтезирует две РНК-затравки. Образуются две репликативные вилки (2 лидирующие и 2 отстающие цепи), которые в процессе комплементарного синтеза удаляются друг от друга, двигаясь в разных направлениях. Наблюдается образование тета-молекул (θ);
3) сбрасывание внутримолекулярного напряжения обеспечивает топоизомераза I путем внесения точечных однонитевых разрывов, которые тут же лигируются. Образуются два двунитевых кольца, где родительские цепи соединены друг с другом. Разъединение осуществляет топоизомераза II, которая вносит разрывы в двунитевые кольца. Затем разрывы лигируются. Такой тип репликации используют в качестве промежуточной стадии многие крупные ДНК-содержащие вирусы, в том числе: бактриофаги, вирусы герпеса, а также, вирусы с кольцевым днДНК геномом, поражающие человека и животных. Это вирусы, относящиеся к семействам Polyomaviridae (см. вирион SV40) и Papilomaviridae, ранее входившие в одно семейство Papovaviridae. Полиома- и папиломавирусы – это б/о, относительно мелкие (4555 нм) икосаэдрические вирусы. Капсид, образованный тремя белками, имеет четко выраженную капсомерную структуру (72 капсомера).
Реплицируются в ядре. Геном – двунитевая кольцевая сверхспирализованная ДНК размером 5-8 т.п.н., ассоциированная с 4-мя клеточными гистонами. Кодирует два неструктурных белка – большой и малый Т-АГ. Это трансформирующие антигены. Геном может интегрировать с геномом клетки хозяина. Большой Т-АГ обладает свойством хеликазы и принимает участие в репликации ДНК – связывается с ДНК в точке ori. Гены Т-АГ транскрибируются сразу после попадания ДНК в ядро. Т.о. транскрипция ДНК у этих вирусов опережает репликацию.
Читать дальшеИнтервал:
Закладка:









