Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Жанр:
- Издательство:Литагент БИБКОМ
- Год:2012
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных краткое содержание
Общая вирусология с основами таксономии вирусов позвоночных - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
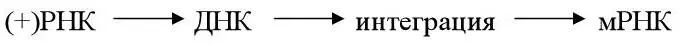
4 днРНК-геном реовирусов. Транскрипцию осуществляет вирионная транскриптаза – РНК-зависимая РНК-полимераза, находящаяся в составе однокапсидной вирусной частицы.
У сложно устроенных РНК-содержащих вирусов транскрипция происходит не на голой матрице, а в составе транскрипционных комплексов. Капсидные белки обеспечивают правильную конформацию РНК, защиту от клеточных нуклеаз, связь фрагментов генома друг с другом, а также регуляцию транскрипции.

5 Амбисенс (обоюдозначащая, амбиполярная) РНК. Амбиполярная РНК имеет 5'-конец позитивной полярности, а 3'-конец негативной полярности. При попадании в клетку с геномной РНК считываются два класса комплементарных РНК: с 3'-конца считывается субгеномная мРНК и полноразмерный антигеном. Антигеном, в свою очередь, опять служит матрицей для двух классов РНК. С антигенома считывается вторая субгеномная мРНК и полноразмерная обоюдозначащая геномная, которая идет на построение вирусных частиц.
3.7.2.3 Особенности транскрипции ДНК-геномов вирусов
Каждая клетка млекопитающих содержит приблизительно 6×10 9пар азотистых оснований ДНК и > 10 4активных промотора. Это создает определенные трудности для вирусной ДНК, т.к. вирусные промоторы должны эффективно конкурировать за использование клеточного транскрипционного аппарата, а завербованный аппарат клетки должен эффективно генерировать вирусные транскрипты. Для этого ДНК-вирусы осуществляют стратегию эффективного инициирования транскрипции на вирусной ДНК в одном или более специфическом промоторе вскоре после того, как ДНК введена в ядро.
Эти промоторы непосредственно управляют транскрипцией так называемых ранних генов.
Энхансеры (гены – усилители). Для того чтобы элементы клеточного транскрипционного аппарата могли взаимодействовать с ранними вирусными промоторами и активизировать транскрипцию, многие ДНК-вирусы содержат геныусилители или энхансеры – cis-acting регуляторные последовательности, выполняющие роль минимальных промоторов.
В отличие от промоторов, они могут активизировать транскрипцию, находясь выше или ниже сайта связывания, запускающего транскрипцию, со скоростью от тысячи пар азотистых оснований (биты) в секунду. Эта особенность определенных энхансеров подчеркивает конкурентоспособное преимущество к связыванию с вирусными промоторами. Действительно, высокая активность энхансера/промотора ранних генов цитомегаловируса определяет его частое использование в векторах экспрессии. Хотя клеточные гены часто содержат энхансеры, первый ген-усилитель был обнаружен у полиомавируса SV40. Энхансер SV40 состоит из двух тандемных дупликаций 72 п.н., которые содержат сайты связывания, по крайней мере, для шести различных факторов транскрипции, включая множественные сайты связывания для некоторых факторов. Необходимым условием для оптимальной функции энхансера является его специфическое местоположение и кратность факторов, участвующих в транскрипции.
Многие энхансеры также содержат специфические сайты для "архитектурных" белков, которые не имеют функции активации транскрипции, а образуют с энхансером собственный, скорее стереоспецифический, трехмерный комплекс белка и гена, который назван энхансеосомой. Эти комплексы нуклеопротеида могут активировать транскрипцию со значительной мощностью и специфичностью за счет взаимодействия с РНК-полимеразой II.
Факторы транскрипции вириона. В дополнение к энхансерам некоторые ДНК-вирусы кодируют белки, которые упакованы в вирион, вводятся в клетку с вирусной ДНК и, затем, облегчают транскрипцию ранних генов. Классическим примером этого являются поксвирусы, которые реплицируются в цитоплазме, что делает бесполезным клеточный аппарат транскрипции. Поксвирусы кодируют и упаковывают в вирионы собственную РНК-полимеразу, ферменты кэппирования и полиаденилирования и факторы транскрипции. После проникновения в клетку и раздевания, вирусный транскрипционный аппарат активируется и синтезирует вирусные ранние транскрипты. Другой примером является трансактиватор VP16 вируса простого герпеса (HSV). HSV VP16 синтезируется на поздней стадии инфекции и упаковывается в часть вириона, известную как тегумент, который находится между капсидом и оболочкой. После проникновения HSV в клетку, VP16 транспортируется в ядро, где формирует комплекс, по крайней мере, с двумя клеточными белками – Oct-1, который является активатором транскрипции, узнающим 8 п.н. последовательность ДНК, и HCF-1. Этот трехмерный комплекс связывается со специфической ДНКпоследовательностью HSV в области TAATGARAT (R = пурин), расположенной выше ранних генов. Специфичность и аффинное сродство обеспечивает Oct-1. Комплекс служит мощным активатором транскрипции. Эта функция связана с С- терминальным активационным доменом VP16, который может связываться фактически с любой последовательностью ДНК, расположенной выше TATA-бокса промотора, в связи с чем, он может активизировать транскрипцию в любой клетке эукариот. Действительно, VP16 является универсальным активатором транскрипции, т.к. активационный домен VP16 может взаимодействовать со многими различными белками клеточного аппарата транскрипции, однако не ясно, какой именно белок наиболее важен для его действия.
Стимуляция генной экспрессии вирусными ранними белками. В процессе внутриклеточного жизненного цикла ДНК-содержащие вирусы экспрессируют свои гены в высоко упорядоченной последовательности. Хотя экспрессия геномов вирусов может регулироваться посттранскрипционно, главный контроль осуществляется на уровне транскрипции. Вирусные ранние гены кодируют белки, которые, будучи транслированы, стимулируют экспрессию так называемых поздних вирусных генов. Это происходит благодаря наличию ранних и поздних промоторов, которые обычно испытывают недостаток энхансеров или специфических сайтов для комплексов транскрипции, подобных тем, что формируются белками вириона или, например, VP16, для того, чтобы эффективно конкурировать с клеточными промоторами транскрипции. Выражение некоторых вирусных генов (например, поздних генов папиломавирусов) ограничено специфическим состоянием клеточной дифференцировки. В основе этого ограничения может лежать ткане-специфическая экспрессия клеточных факторов транскрипции.
Должно также быть отмечено, что несколько вирусов кодируют белки, которые дополнительно стимулируют экспрессию некоторых вирусных генов в специфическое время жизненного цикла вируса. Кроме того, многие вирусы влияют на экспрессию клеточных генов, что может повлечь за собой индукцию или репрессию специфических генов, а также неспецифическое выключение экспрессии генов. Имеется много механизмов, которые связаны с функцией ранних вирусных белков. Например, белок BZLF1 (также известный, как Zta) вируса Эпштейна-Барр (сем. Herpesviridae) , может вести себя подобно клеточным факторам транскрипции, которые связываются со специфической последовательностью выше генов и активизируют их транскрипцию.
Читать дальшеИнтервал:
Закладка:









