Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Жанр:
- Издательство:Литагент БИБКОМ
- Год:2012
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных краткое содержание
Общая вирусология с основами таксономии вирусов позвоночных - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Другие ранние белки увеличивают активность клеточных факторов транскрипции. Например, белок аденовируса E1A замещает членов транскрипции, разрушает комплекс белков семейства E2F и белков-репрессоров семейства Rb. Это увеличивает транскрипцию ранних генов аденовируса, которые содержат E2Fсвязывающие сайты, что имеет важные последствия для клеточной транскрипции и, косвенно, для репликации вирусной и клеточной ДНК. Другие ранние белки, подобные ICP4 вируса простого герпеса, могут стимулировать транскрипцию путем прямого взаимодействия с транскрипционным аппаратом без связывания со специфическими последовательностями ДНК целевого промотора, или стимулировать экспрессию вирусных генов с использованием механизмов посттранскрипционного процессинга. В любом случае результатом действий этих вирусных белков является то, что клеточный транскрипционный аппарат используется для реализации информационных программ вируса, а не клетки.
Принципы транскрипции ДНК-геномов. По механизму транскрипции ДНК-геномы вирусов можно разделить на 4 группы:
1 днДНК, транскрипцию которой осуществляют клеточные ферменты (ядерные полиома-, адено-, герпесвирусы, ряд бактериофагов, например λ, Т4). Как правило, транскрипцию осуществляет РНК-полимераза II, реже – РНК-полимераза III (ряд транскриптов аденовируса). Возможны ситуации, когда транскрипцию ранних генов осуществляет клеточная РНК-полимераза, а поздних – вирусоспецифическая или модифицированная клеточная РНК-полимераза, содержащая вирусоспецифическую субъединицу (фаги Т7 и Т4, соответственно).

2 днДНК, раннюю транскрипцию которой осуществляют вирусные полимеразы, входящие в состав вириона. Поздние гены транскрибируются вновь синтезированными вирусными полимеразами, детерминированными геномом вируса (поксвирусы, фаг N4).

3 онДНК, которая прежде чем транскрибироваться должна перейти в двухнитевую форму (парвовирусы, включая дефектные вирусы, требующие для размножения наличия вируса-помощника).
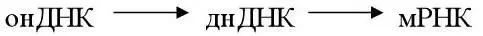
4 Кольцевая частично двухнитевая днДНК (вирус гепатита B, вирус мозаики цветной капусты). Прежде чем транскрибироваться, ДНК проходит стадию репарации.
3.7.2.4 Регуляция транскрипции
В вирусных и клеточных системах молекулярные механизмы транскрипции принципиально сходны. Отличие заключается в существовании различных способов регуляции транскрипции вирусных геномов. Необходимость такой регуляции определяется разной потребностью в вирусоспецифических белках. Структурные белки, как правило, требуются в больших количествах, чем белки-ферменты. Кроме того, на ранних стадиях инфекции нужны белки, обеспечивающие репликацию вирусного генома, а на поздних – структурные белки. Поэтому целесообразно, чтобы разные вирусные гены считывались с разной эффективностью, и эта эффективность менялась во времени.
Процесс транскрипции регулируется на уровне транскриптона (оперона) за счет работы репрессоров и активаторов белковой природы и энхансеров (усилителей), которые представляют собой определенные короткие последовательности геномной нуклеиновой кислоты. Транскрипция регулируется количественно и качественно и осуществляется как клеточными, так и вирус-специфическими механизмами.
У вирусов установлено существование целого ряда способов регуляции транскрипции.
Временной тип регуляции. У ДНК-содержащих вирусов существует три периода транскрипции: сверхранний, ранний и поздний. При сверхранней и ранней транскрипции считываются сверхранние и ранние гены, при поздней – поздние гены. Количество транскриптов поздних генов превышает количество ранних. Многие сверхранние мРНК являются генами NS белков-ферментов и регуляторов транскрипции и репликации. Поздние мРНК являются генами структурных белков. Фактором регуляции транскрипции у ядерных вирусов является транспорт мРНК в цитоплазму.
Каскадный тип регуляции транскрипции генов. Суть такой регуляции заключается в том, что продукты сверхранней транскрипции, например, α-белки, необходимы для транскрипции другой группы генов, кодирующих β-белки, которые, в свою очередь, включают транскрипцию следующей группы генов – γ-белков.
Полярный тип регуляции определяется порядком расположения генов в геноме. Количество синтезируемых молекул полипептида зависит от расстояния между геном и промотором. Вдоль генома (-)РНК вирусов существует как бы градиент эффективности транскрипции. Чаще транскрибируются гены 3'-региона, реже – гены 5'-конца.
Взаимное расположение и сила регуляторных сигналов. Считывание или несчитывание транскрибируемого участка матрицы зависит от свойств и расположения регуляторных сигналов – промоторов (обеспечивают начало транскрипции) и терминаторов (обеспечивают прекращение транскрипции). Основа регуляции – взаимное расположение регуляторных сигналов и их сила. Активность сигналов может меняться во времени.
Характер образования транскриптов и способ регуляции зависят от того, имеем ли мы дело с вирусами прокариот или эукариот. Напомним, что в клетках прокариот возможна множественная инициация трансляции на полицистронной матрице, тогда как в клетках эукариот на РНК реализуется только одна точка инициации трансляции и эта мРНК функционально моноцистронна. Ограничения, накладываемые клеткой хозяина, в первую очередь сказываются на механизмах транскрипции и посттранскрипционного созревания мРНК. Приведем конкретные примеры способов регуляции транскрипции вирусных геномов в клетках прокариот и эукариот.
Самый простой способ регуляции транскрипции в клетках прокариот установлен у фагов 1М13 и fd, где разная степень экспрессии фаговых генов регулируется за счет расположения и силы промоторов. За счет наличия «сильных» промоторов активно транскрибируются гены, кодирующие основной структурный белок капсида и ДНК-связывающий белок. В то же время, геномная последовательность, кодирующая минорные вирусные белки, имеет промоторы, отнесенные к разряду «слабых».
Более сложная регуляция транскрипции генов наблюдается у фага λ, имеющего, как минимум, три типа регуляции транскрипции: 1) ретро-регуляция – осуществляется при участии нуклеотидных последовательностей, расположенных за транскрибируемым геном. Этот участок комплементарен предшествующему участку гена и в образовавшемся транскрипте возникает внутримолекулярная двухнитевая структура, которая впоследствии разрушается РНКазой III; 2) аутогенная регуляция – регуляция активности гена при помощи продукта этого же гена; 3) индукция профага – наблюдается в результате инактивации репрессора.
Читать дальшеИнтервал:
Закладка:









