Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Жанр:
- Издательство:Литагент БИБКОМ
- Год:2012
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных краткое содержание
Общая вирусология с основами таксономии вирусов позвоночных - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Негерметичное сканирование при инициации трансляции популярно среди вирусов, а у ретровирусов оно может обеспечивать определенные стехиометрические взаимоотношения продуктов трансляции.
Удлинение. В течение стадии элонгации трансляции мРНК связана со многими 80S рибосомами, или полисомами, поскольку аминокислотные остатки последовательно присоединяются к СООН-концу растущей цепи пептидов. Во многих вирусных системах жизненный цикл разграничен на ранние и поздние события, которые могут различаться дифференцированным привлечением вирусных мРНК в полисомные комплексы в определенное время после инфекции. Например, у вируса простого герпеса (HSV-1) это часто совпадает с синтезом факторов латентности и детерминант вирулентности. Процесс трансляции на стадии элонгации подчинен вирусной регуляции. Механизмы контроля элонгации включают, например, рибосомальный сдвиг рамки считывания и направленную вирусом модификацию EF-1. Первый механизм распространен у ретровирусов и связан с наличием дополнительных ORFs в пределах вирусной мРНК.
Терминация. Процесс завершения трансляции происходит в тот момент, когда 80S рибосома сталкивается в рамке считывания с терминирующим кодоном в пределах последовательности мРНК. Терминирующий кодон является фактором, который запускает процесс гидролиза связи пептидной цепи и тРНК, освобождает синтезированный полипептид от 80S рибосомы и разобщает субъединицы рибосомы. Как только завершение синтеза произошло, 40S субъединица может продолжить сканировать мРНК.
При считывании мультицистронной последовательности завершение трансляции может сопровождаться переинициированием трансляции ниже расположенного гена. Завершение трансляции переинициированием распространено среди вирусов и используется ими, чтобы управлять синтезом определенного генного продукта. После открытия полиаденилирования э-мРНК стало ясно, что поли-A трек играет важную роль в трансляции мРНК в клетках эукариот. Современные исследования показали, что определенную роль в стимулирующей функции поли-A трека на процесс трансляции играет поли-A-связывающий белок (PABP). В клетках животных PABP взаимодействует с элементами кэп-связывающего комплекса, включая в его состав 5’– конец мРНК и создавая, таким образом, трансляционный комплекс в форме «закрытой петли» (рисунок 17).
мРНК связующий EIF4F инициирующий комплекс взаимодействует с 3’– концом мРНК через PABP. Поли-A последовательность в пределах 3’-НТР прямым образом связывает PABP с мРНК. PABP добивается взаимодействия с кэпсвязывающим комплексом непосредственно через eIF4G (4G) или косвенно через взаимодействие eIF4G, eIF4A (4A) и Paip-1. Сборка комплекса замкнутой системы может стабилизировать взаимодействие 40S субъединицы рибосомы с мРНК.
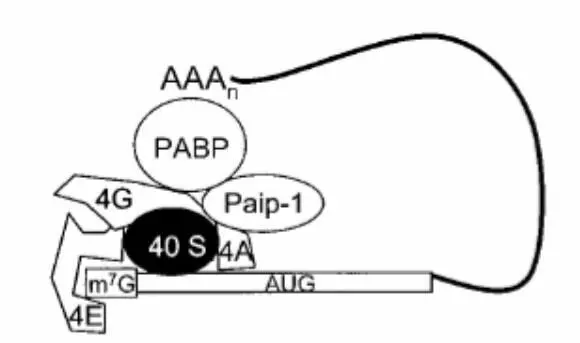
Рисунок 17 – Модель трансляционного комплекса в замкнутой системе с мРНК
PABP формирует закрытую петлю путем связывания eIF4G и белка Paip-1. Paip-1 взаимодействует с компонентами кэп-связывающего комплекса мРНК, включая eIF4G и eIF4A-хеликазу. Изучение инициации трансляции в дрожжах и растениях показали, что взаимодействие между PABP и eIF4G стимулирует трансляцию мРНК. Сближение концов мРНК, обеспеченное закрытым трансляционным комплексом, вносит вклад в стабильность мРНК и 5’-кэп-комплекса и обеспечивает эффективную сборку полирибосом. Таким образом, полный эффект закрытой петли заключается в увеличении эффективности трансляции. Вирусы используют закрытый трансляционный комплекс как средство переключения трансляционного аппарата клетки на трансляцию вирусных мРНК путем разрушения или модификации РАВР.
4 Репродукция вирусов
Процесс репродукции вирусов может быть условно разделен на две фазы. Первая фаза охватывает события, которые ведут к адсорбции и проникновению вируса в клетку, освобождению его внутреннего компонента и модификации его таким образом, что он способен вызвать инфекцию. Соответственно, первая фаза включает в себя три стадии:
1) адсорбция вируса на клетках;
2) проникновение в клетки;
3) раздевание вируса в клетке.
Эти стадии направлены на то, чтобы вирус был доставлен в соответствующие клеточные структуры, и его внутренний компонент был освобожден от защитных оболочек. Как только эта цель достигнута, начинается вторая фаза репродукции, в течение которой происходит экспрессия вирусного генома. Эта фаза включает в себя стадии:
1) транскрипции;
2) трансляции информационных РНК;
3) репликации генома;
4) сборки вирусных компонентов. Заключительной стадией репродукции является выход вируса из клетки.
4.1 Адсорбция
Взаимодействие вируса с клеткой начинается с процесса адсорбции, т.е. прикрепления вирусных частиц к клеточной поверхности. Процесс адсорбции возможен при наличии соответствующих рецепторов на поверхности клетки и «узнающих» их субстанций на поверхности вируса. Самые начальные процессы адсорбции имеют неспецифический характер, и в основе их может лежать электростатическое взаимодействие положительно и отрицательно заряженных группировок на поверхности вируса и клетки. Однако узнавание клеточных рецепторов вирусными белками, ведущее к прикреплению вирусной частицы к клетке, является высоко специфическим процессом. Белки на поверхности вируса, узнающие специфические группировки на плазматической мембране клетки и обусловливающие прикрепление к ним вирусной частицы, называются прикрепительными белками (рисунок 18).
Вирусы используют рецепторы, предназначенные для прохождения в клетку необходимых для ее жизнедеятельности веществ: питательных веществ, гормонов, факторов роста и т.д. Рецепторы могут иметь разную химическую природу и представлять собой белки, углеводный компонент белков и липидов, липиды. Рецепторами для вирусов гриппа и парамиксовирусов является сиаловая кислота в составе гликопротеидов и гликолипидов (ганглиозидов), для рабдовирусов и реовирусов – также углеводный компонент в составе белков и липидов, для пикорна- и аденовирусов – белки, для некоторых вирусов – липиды. Специфические рецепторы играют роль не только в прикреплении вирусной частицы к клеточной поверхности. Они определяют дальнейшую судьбу вирусной частицы, ее внутриклеточный транспорт и доставку в определенные участки цитоплазмы и ядра, где вирус способен инициировать инфекционный процесс. Вирус может прикрепиться и к неспецифическим рецепторам и даже проникнуть в клетку, однако только прикрепление к специфическому рецептору приведет к возникновению инфекции.
Читать дальшеИнтервал:
Закладка:









