Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Жанр:
- Издательство:Литагент БИБКОМ
- Год:2012
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных краткое содержание
Общая вирусология с основами таксономии вирусов позвоночных - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
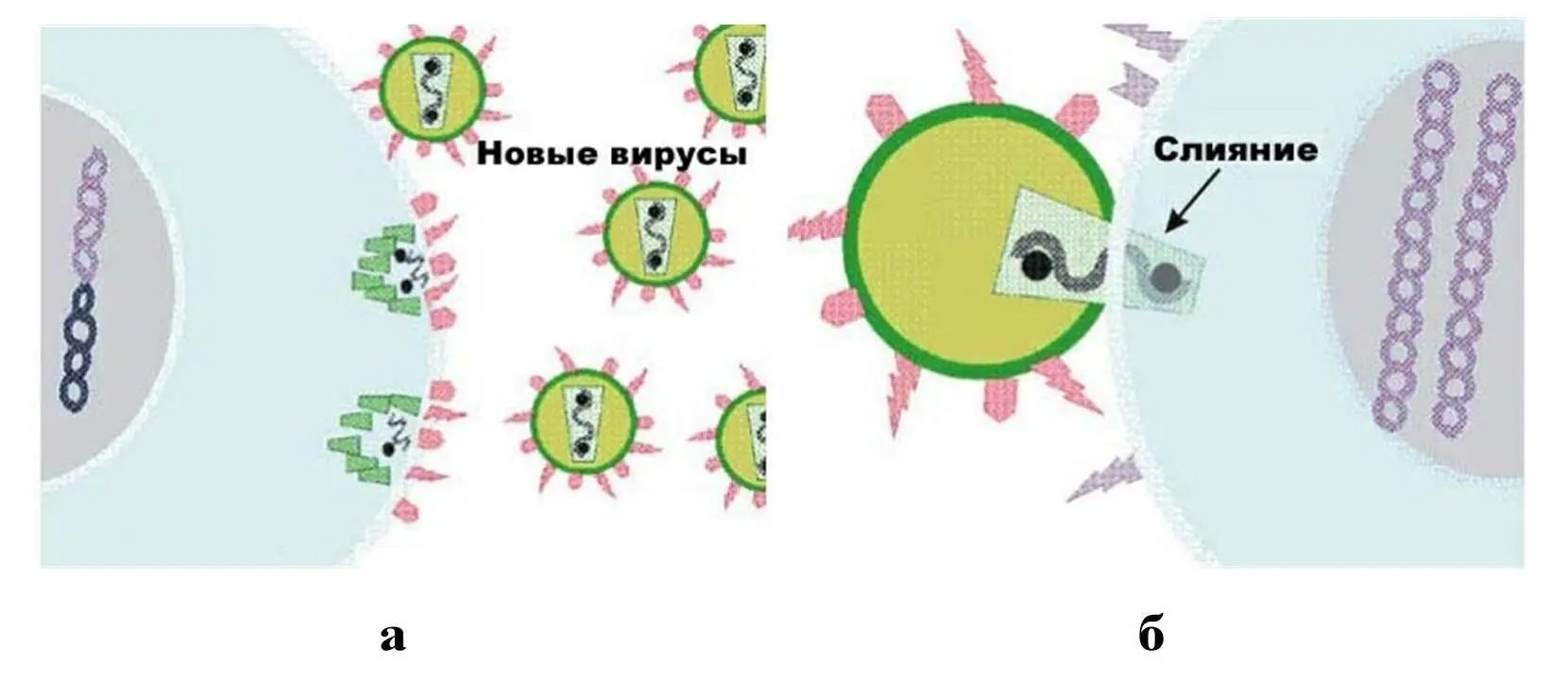
а – узнавание клеточных рецепторов вирусными белками, ведущее к прикреплению вирусной частицы к клетке; б – прикрепление вируса к клетке.
Рисунок 18 – Адсорбция вируса на клетке
Прикрепление вирусной частицы к клеточной поверхности вначале происходят путем образования единичной связи вирусной частицы с рецептором. Однако такое прикрепление непрочно, и вирусная частица может легко оторваться от клеточной поверхности (обратимая адсорбция). Для того чтобы наступила необратимая адсорбция, должны появиться множественные связи между вирусной частицей и многими молекулами рецепторов, т.е. должно произойти стабильное мультивалентное прикрепление. Количество молекул клеточных рецепторов в участках адсорбции может доходить до 3000. Стабильное связывание вирусной частицы с клеточной поверхностью в результате мультивалентного прикрепления происходит благодаря возможности свободного перемещения молекул рецепторов в липидном бислое плазматической мембраны, которое определяется подвижностью, «текучестью» белково-липидного слоя. Увеличение текучести липидов является одним из наиболее ранних событий при взаимодействии вируса с клеткой, следствием которого является формирование рецепторных полей в месте контакта вируса с клеточной поверхностью и стабильное прикрепление вирусной частицы к возникшим группировкам – необратимая адсорбция.
Количество специфических рецепторов на поверхности клетки колеблется между 10 4и 10 5на одну клетку. Рецепторы ряда вирусов могут быть представлены лишь в ограниченном наборе клеток-хозяев, и этим может определяться чувствительность организма к данному вирусу. Например, пикорнавирусы адсорбируются только на клетках приматов. Рецепторы для других вирусов, напротив, широко представлены на поверхности клеток различных видов, как, например, рецепторы для ортомиксовирусов и парамиксовирусов, представляющие собой сиалилсодержащие соединения. Поэтому эти вирусы имеют относительно широкий диапазон клеток, на которых может происходить адсорбция вирусных частиц. Рецепторами для ряда тогавирусов обладают клетки исключительно широкого круга хозяев: эти вирусы могут адсорбироваться к инфицировать клетки, как позвоночных, так и беспозвоночных.
Наличие специфических рецепторов на поверхности клетки в ряде случаев обусловливает феномен зависимого от хозяина ограничения, т.е. способность вируса заражать лишь определенные виды животных. В целом ограничения при взаимодействии рецепторных систем вируса и клетки биологически оправданы и целесообразны, хотя в ряде случаев они являются «перестраховкой». Так, многие линии клеток, устойчивых к вирусам полиомиелита и Коксаки, можно заразить депротеинизированными препаратами РНК, выделенными из этих вирусов. Такое заражение клеток идет в обход естественных входных путей инфекции через взаимодействие с клеточными рецепторами. Известна потенциальная способность вирусов животных реплицироваться в протопластах дрожжей, грибов и бактерий, а бактериофагов – в клетках животных. Таким образом, вирусные ДНК и РНК обладают способностью заражать и более широкий круг хозяев, чем вирусы.
Вирусные прикрепительные белки. Прикрепительные белки могут находиться в составе уникальных органелл, таких как структуры отростка, у Т-бактериофагов или фибры у аденовирусов, которые хорошо видны в электронном микроскопе; могут формировать морфологически менее выраженные, но не менее уникальные аранжировки белковых субъединиц на поверхности вирусных мембран, как, например, шипы у оболочечных вирусов, «корону» у коронавирусов.
Просто организованные вирусы животных содержат прикрепительные белки в составе капсида. У сложно организованных вирусов эти белки входят в состав суперкапсида и представлены множественными молекулами. Например, у вируса леса Семлики (альфа-вирус) имеется 240 молекул гликопротеида в одном вирионе, у вируса гриппа – 300-450 гемагглютинирующих субъединиц, у реовируса – 24 молекулы белка, у аденовируса – 12 фибров.
4.2 Проникновение вирусов в клетку
Исторически сложилось представление о двух альтернативных механизмах проникновения в клетку вирусов животных – путем виропексиса (эндоцитоза) и путем слияния вирусной и клеточной мембран (рисунок 19). Однако оба эти механизма не исключают, а дополняют друг друга.
Термин «виропексис», предложенный в 1948 г. Фазекасом де сан Гро, означает, что вирусная частица попадает в цитоплазму в результате инвагинации участка плазматической мембраны и образования вакуоли, которая содержит вирусную частицу.
Рецепторный эндоцитоз. Виропексис представляет собой частный случай рецепторного или адсорбционного эндоцитоза. Этот процесс является обычным механизмом, благодаря которому в клетку поступают питательные и регуляторные белки, гормоны, липопротеины и другие вещества из внеклеточной жидкости. Рецепторный эндоцитоз происходит в специализированных участках плазматической мембраны, где имеются специальные ямки, покрытые со стороны цитоплазмы особым белком с большой молекулярной массой – клатрином. На дне ямки располагаются специфические рецепторы. Ямки обеспечивают быструю инвагинацию и образование покрытых клатрином внутриклеточных вакуолей. Полупериод проникновения вещества внутрь клетки по этому механизму не превышает 10 мин с момента адсорбции. Количество образующихся в одну минуту вакуолей достигает более 2000. Таким образом, рецепторный эндоцитоз представляет собой хорошо слаженный механизм, который обеспечивает быстрое проникновение в клетку чужеродных веществ.
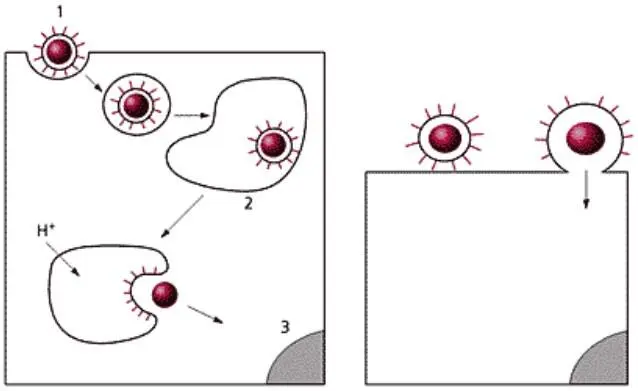
а – виропексис (эндоцитоза); б – слияние вирусной и клеточной мембран.
Рисунок 19 – Механизмы проникновения в клетку вирусов
Покрытые вакуоли сливаются с другими, более крупными цитоплазматическими вакуолями, образуя рецептосомы, содержащие рецепторы, но не содержащие клатрин, а те в свою очередь сливаются с лизосомами. Таким путем проникшие в клетку белки обычно транспортируются в лизосомы, где происходит их распад на аминокислоты; они могут и миновать лизосомы, и накапливаться в других участках клетки в недеградированной форме. Альтернативой рецепторного эндоцитоза является жидкостный эндоцитоз, когда инвагинация происходит не в специализированных участках мембраны.
Большинство оболочечных и безоболочечных вирусов животных проникает в клетку по механизму рецепторного эндоцитоза. Эндоцитоз обеспечивает внутриклеточный транспорт вирусной частицы в составе эндоцитарной вакуоли, поскольку вакуоль может двигаться в любом направлении и сливаться с клеточными мембранами (включая ядерную мембрану), освобождая вирусную частицу в соответствующих внутриклеточных участках. Таким путем, например, ядерные вирусы попадают в ядро, а реовирусы – в лизосомы. Однако проникшие в клетку вирусные частицы находятся в составе вакуоли и отделены от цитоплазмы ее стенками. Им предстоит пройти ряд этапов, прежде чем они смогут вызвать инфекционный процесс.
Читать дальшеИнтервал:
Закладка:









