Борис Стрельцов - Хранение цветов

- Название:Хранение цветов
- Автор:
- Жанр:
- Издательство:Агропромиздат
- Год:1988
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Борис Стрельцов - Хранение цветов краткое содержание
Хранение цветов - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Проникновение кислорода в глубь растительных тканей зависит и от морфологических свойств органов растений. В основном газообмен срезки происходит через устьица, на внешний покров — кутикулу приходится лишь небольшая его часть. Там, где отсутствуют устьица, например у семян, луковиц, газообмен идет через толстую кутикулярную оболочку постепенно и зависимость поглощения кислорода имеет вид прямой. Отклонения от прямой линии в сторону большего (гиперболического) роста интенсивности дыхания с повышением концентрации кислорода свидетельствуют о том, что, начиная с некоторых пороговых величин, концентрация кислорода перестает быть единственным фактором, ограничивающим дыхание, и начинает проявляться влияние других факторов: концентрации дыхательных субстратов, активности ферментного аппарата и т. п. Кроме того, передвижение кислорода в растительном организме может происходить не только диффузионным путем.
Для проведения расчетов, связанных с установлением газового режима в хранилищах, контейнерах, упаковках с цветочной продукцией, в особенности для режимов, характеризующихся относительно малой концентрацией кислорода (3–5 °C), можно принять линейную зависимость интенсивности дыхания [м 3/(кгс)] от концентрации кислорода:
К — K 0V 1/V, (3)
где K 0— интенсивность дыхания в обычной атмосфере (21 % O 2), м 3/кгс; V 1, V — соответственно первоначальный и текущий объемы кислорода, м 3.
Выделяющийся в процессе дыхания углекислый газ также оказывает определенное влияние на интенсивность дыхания. Возрастание концентрации этого газа приводит к некоторому снижению интенсивности дыхания, но этот эффект проявляется лишь при относительно больших значениях содержания СO 2. Как углекислый газ, так и различные углеводороды блокируют активность ферментов биологического окисления.
На процессы дыхания в значительной степени влияет и водный режим растений. Интенсивность дыхания созревающих семян уменьшается по мере уменьшения их влагосодержания, и сухие семена, луковицы, клубнелуковицы дышат относительно слабо, так как находятся в состоянии физиологического покоя. Добавление влаги выводит их из этого состояния, что сопровождается возрастанием интенсивности дыхания. Иная картина наблюдается при подсыхании срезки цветов. Сначала интенсивность дыхания повышается, после чего она снижается до уровня более низкого, чем вначале. Если потеря влаги не вызвала необратимого повреждения тканей цветка, то дыхание может стабилизироваться на таком более низком уровне. Если затем срезка будет обеспечена достаточным количеством воды, то дыхание может возвратиться к исходному уровню, при условии что пониженная влажность, которая часто бывает при так называемом сухом хранении, не привела к падению влагосодержания ниже критической величины. В противном случае возобновление водоснабжения уже не восстанавливает нормальное дыхание, происходит дальнейшее разрушение клеточных структур, и растение гибнет.
При изменении содержания влаги дыхательный коэффициент может изменяться в разные стороны, что связывают с неодинаковой чувствительностью различных ферментов к влаге и с возможными субмикроскопическими изменениями клеточных структур, определяющими диффузию дыхательных субстратов. Помимо этого, влагосодержание влияет на проницаемость клеточных мембран для разных газовых компонентов.
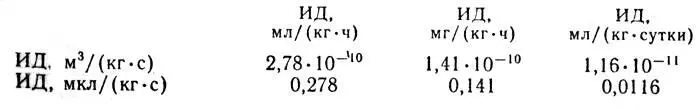
Рассмотрим пример пересчета. Известно, что лепестки розы имеют интенсивность дыхания 500 мг СО 2/(кгч). Необходимо выразить эту величину в м 3/(кг•с). С учетом приведенного выше соотношения получим: К = 1,41•10 -10•500 =7,05•10 -8, м 3/(кг•с). Это значение соответствует также 70,5 мкл/(кг•с). В таблице 2 приведены данные об интенсивности дыхания черенков и срезки некоторых цветов.
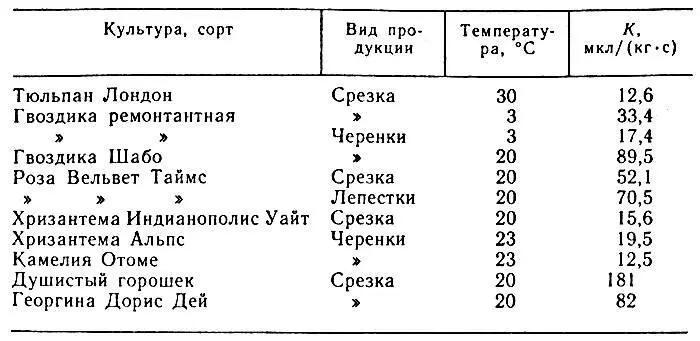
Таблица 2. Интенсивность дыхания цветов и черенков
ТРАНСПИРАЦИЯ
Вода — главный компонент живых растительных клеток, на ее долю приходится до 90 % сырой массы. В зрелых растительных клетках большая часть воды содержится в крупной центральной вакуоли (Гэлстон, 1983), которая плотно прижимается к клеточной стенке, способствуя поддержанию формы клетки в мягких органах растения (листьях, стеблях, побегах). Форма декоративных травянистых растений зависит от гидростатических сил, при ослаблении последних вследствие потери влаги возникают утрата тургора и увядание. Если процесс увядания, например при сухом хранении срезки, не зашел далеко, при получении влаги запас воды в вакуоли возобновляется, тургор восстанавливается. Растения за счет специальных механизмов саморегуляции способны поддерживать свою оводненность на постоянном уровне, непрерывно поглощая воду и испаряя ее.
Транспирация представляет собой выделение растением водяного пара в атмосферу в основном посредством листьев и лепестков. Вода испаряется с поверхности влажных клеток, диффундирует по межклетникам и выходит наружу через открытые устьица либо кутикулу. Способность воды диффундировать, испаряться или поглощаться оценивают величиной водного потенциала, имеющего размерность давления. В отсутствие противодавления клеточной стенки поступление воды в клетку определяется водным потенциалом клетки, первоначально равным осмотическому потенциалу раствора, заполняющего вакуоль. С проникновением воды в вакуоль объем последней увеличивается, а раствор становится более разбавленным. Цитоплазма прижимается к стенке клетки, возникает противодавление клеточной стенки на клеточное содержимое. Когда противодавление клеточной стенки достигает достаточно большой величины, устанавливается динамическое равновесие, и приток воды прекращается.
Водный потенциал клетки определяется суммой потенциала давления и осмотического потенциала, который всегда отрицателен. Потенциал давления в живых клетках положителен. Клеточный потенциал равен нулю (при полном тургоре) или отрицателен при потере влаги и тургора. Если потенциал давления или тургорный потенциал упал до нуля, лист полностью завядает. Поэтому о завядании и потере влаги листьями и лепестками обычно судят по легко измеряемой величине водного потенциала соответствующих клеток (см. главу 8).
Интенсивность транспирации зависит от температуры и влажности воздуха, ширины раскрытия устьичных щелей, числа устьиц, облиственности срезки и разности водных потенциалов воздуха внутри и снаружи листа. Обычно вода из межклетников диффундирует в более сухой атмосферный воздух, и продолжительность сохранения срезки цветов в свежем состоянии определяется тем, как обеспечен транспорт к цветку воды и метаболитов, необходимых для поддержания жизнедеятельности.
Читать дальшеИнтервал:
Закладка:








