Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
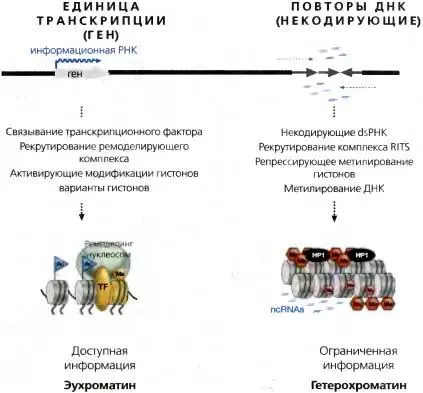
Рис. 3.9.Различие между эухроматиновыми и гетерохроматиновыми доменами
Рисунок, суммирующий различия между эухроматином и конститутивным гетерохроматином. В их числе — различия в типе продуцируемых транскриптов, в рекрутировании связывающихся с ДНК белков (т. е. транскрипционного фактора [TF]), в ассоциированных с хроматином белках и комплексах, в ковалентных модификациях гистонов и в составе вариантных гистонов
7. Модификации гистонов и гистоновый код
Мы рассмотрели, каким образом модификации гистонов могут изменять хроматиновую матрицу посредством cis -эффектов, изменяющих межнуклеосомные контакты и промежуточные расстояния, или же trans -эффектов, вызываемых ассоциациями гистоновых и негистоновых белков с матрицей. Каков же вклад и биологический результат модификаций гистонов? Типы структуры хроматина, коррелирующие с модификациями гистоновых «хвостов», выявились в результате исследований, в которых использовалась основная масса гистонов и которые позволили предположить, что эпигенетические метки могут обеспечить сигнатуры «ВКЛЮЧЕНО» (т. е. активно) или «ВЫКЛЮЧЕНО» (т. е. неактивно). Это выяснилось в результате длительных исследований большей частью коррелятивного характера, которые показали, что определенные модификации гистонов, в особенности их ацетилирование, связаны с доменами активного хроматина или с участками, в целом пермиссивными для транскрипции В противоположность этому другие метки, такие как некоторые фосфорилированные остатки гистонов, давно ассоциировались с конденсированным хроматином, который, в целом, не поддерживает транскрипционную активность. Модификации гистонов, приведенные в Приложении 2, суммируют известные в настоящее время сайты модификаций. Здесь мы подчеркиваем, что приведенный материал отражает модификации и сайты, которые не обязательно характерны для любого организма.
Прежде всего, обсудим, каким образом производятся или удаляются модификации гистонов. Большая работа, проведенная в области исследования хроматина, позволила предположить, что модификации «хвостов» гистонов образуются («записываются») или удаляются («стираются») в результате каталитического действия ассоциированных с хроматином ферментативных систем. Однако исследователям годами не удавалось идентифицировать эти ферменты. За последнее десятилетие было идентифицировано очень большое число модифицирующих хроматин энзимов из различных источников; большая часть этих данных собрана в Приложении 2. Этот успех был достигнут в результате многочисленных биохимических и генетических исследований. Эти ферменты часто входят в состав крупных, состоящих из многих субъединиц комплексов, которые могут катализировать включение или удаление ковалентных модификаций как в гистоновых, так и в негистоновых мишенях. Более того, многие из этих энзимов катализируют свои реакции с замечательной специфичностью по отношению к остатку-мишени и к клеточному контексту (т. е. в зависимости от внешних или внутренних сигналов). Для ясности, а также в качестве примера, мы кратко обсудим четыре главные ферментативные системы, которые катализируют модификации гистонов, вместе с ферментативными системами противоположного действия, обеспечивающими реверсию этих модификаций (рис. 3.10) (Vaquero et al., 2003; Holbert and Marmorstein, 2005). В совокупности эти антагонистические активности управляют устойчивым балансом каждой рассматриваемой модификации.
Ацетилазы гистонов (HATs) ацетилируют остатки специфических лизинов в гистоновых субстратах (Roth et al., 2001); реверсия обеспечивается действием деацетилаз гистонов (HDACs) (Grozinger and Schreiber, 2002). Ферменты семейства гистоновых киназ фосфорилируют остатки специфических серинов или треонинов. а фосфатазы (РРТазы) удаляют метки, созданные фосфорилированием. Особенно хорошо известны митотические киназы, такие как циклин-зависимая киназа или киназа «aurora», катализирующая фосфорилирование корового (H3) и линкерного (Н1) гистонов. Менее очевидны в каждом из этих случаев противостоящие РРТазы, действие которых ревертирует результат этого фосфорилирования, когда клетки выходят из митоза.
Были описаны два общих класса метилирующих ферментов: PRMTs (protein arginine methyltransferases — метилтрансферазы аргининов белка), субстратом которых является аргинин (Lee et al., 2005), и HKMTs (histone lysine methyltransferases — метилтрансферазы лизинов гистонов), действующие на остатки лизина (Lachner et al., 2003). Метилирование аргининов косвенным образом ревертируется действием дезиминаз, которые конвертируют метиларгинин (или аргинин) в остаток цитруллина (Bannister and Kouzarides, 2005). Метилированные остатки лизина оказываются химически более стабильными. Было показано, что метилированный лизин существует в моно-, ди- и триметилированном состояниях. Несколько триметилированных остатков на аминоконцах H3 и Н4, по-видимому, обладают возможностью стабильно воспроизводиться в ходе клеточных делений (Lachner et al., 2004), как и метка H4K20mel в имагинальных дисках Drosophila (Reinberg et al., 2004). Недавно была описана лизин-специфичная «деметилаза» (LSD1) как аминооксидаза, способная удалять метилирование H3К4 (Shi et al., 2004). Этот энзим действует путем FSD-зависимой окислительной дестабилизации аминометильной связи, что приводит к образованию немодифипированного лизина и формальдегида. Было показано, что LSD1 действует избирательно в отношении активирующей H3К4 метки, созданной метилированием, и может дестабилизировать только моно- и ди-, но не триметилирование. Эта деметилаза является частью большого репрессивного белкового комплекса, который содержит также HDACs и другие энзимы Другие данные позволяют предполагать, что LSD1 может соединяться в комплекс вместе с рецептором андрогена в локусах-мишенях и деметилирует репрессивную гистоновую метку H3K9me2, внося вклад в транскрипционную активацию (Metzger et al., 2005). Другой класс гистоновых деметилаз, согласно данной ему храктеристике. работает с помощью более мощного механизма, механизма радикальной атаки, известного как гидроксилазы или диоксигеназы (Tsukada et al., 2006). Одна из них дестабилизирует только H3K36me2 (активная метка), но не триметилированное состояние. Эта новая гистоновая деметилаза jumonji (JHDM1) содержит консервативный домен jumonji, которых в геноме млекопитающих известно около 30; это позволяет предполагать, что некоторые из этих энзимов, может быть, могут атаковать и другие остатки, так же как триметильное состояние (Fodor et al., 2006; Whetstine et al., 2006).
Значительный прогресс был достигнут в анализе систем ферментов, управляющих устойчивым балансом этих модификаций, и мы подозреваем, что в этой увлекательной области будут достигнуты гораздо большие успехи. Остается понять, как регулируются эти ферментативные комплексы и как становятся мишенями их физиологически релевантные субстраты и сайты. Кроме того остается неясным, как ковалентные механизмы влияют на эпигенетические явления.
Читать дальшеИнтервал:
Закладка:



