Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
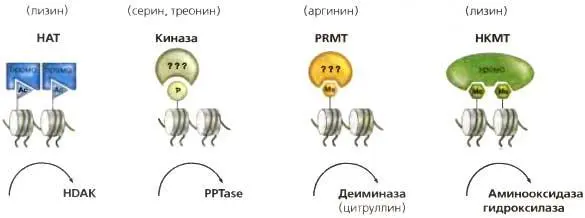
Рис. 3.10.энзимы, модифицирующие гистоны
Ковалентные модификации гистонов трансдуцируются ферментами, модифицирующими гистоны («писателями»), и удаляются активностями противоположного действия. Они делятся на классы в соответствии с типом ферментативного действия (например. ацетилирование или фосфорилирование). Белковые домены со специфическим сродством к модификации гистонового «хвоста» называются «читателями» (HAT) ацетилтрансфераза гистона; (PRMT) протеин-аргинин-метилтрансфераза; (HKMT) метилтрансфераза лизина гистонов; (HDAC) деацетилаза гистонов; (РРТаза) протеинфосфатазы; (Ас) ацетилирование; (Р) фосфорилирование; (Ме) метилирование
Модификации гистонов происходят не изолированно, а на основе комбинирования, как это предполагается — для модификационных кассет (т. е. ковалентных модификаций в соседних остатках определенного гистонового «хвоста», например H3К9те и H3S10ph или H4Slph, H4R3me и H4K4ас) и trans- гистоновых путей (ковалентных модификаций между разными «хвостами» гистонов или нуклеосомами; рис. 3.11). Интригует тот факт что почти все известные модификации гистонов коррелируют с активирующей или репрессивной функциями, в зависимости от того, какой аминокислотный остаток (остатки) в аминоконце гистона модифицирован Описаны как синергистические, так и антагонистические пути (Zhang and Reinberg, 2001; Berger, 2002; Fischle et al., 2003b), которые могут постепенно индуцировать комбинации активных меток, одновременно противодействуя репрессивным модификациям. Неизвестно, однако, сколько разных комбинаций модификаций по различным аминотерминальным позициям гистонов существует для любой данной нуклеосомы, потому что большинство исследований было выполнено на препаратах суммарных гистонов. Недавно было показано, что на структуру и сборку хроматина влияют, кроме аминоконцов, модификации в глобулярных доменах гистонов (Cosgrove et al., 2004), регулируя тем самым экспрессию генов и репарацию повреждений ДНК (van Attikum and Gasser, 2005; Vidanes et al., 2005). Стоит также упомянуть, что несколько модифицирующих гистоны энзимов избирают своей мишенью и негистоновые субстраты (Sterner and Berger, 2000; Chuikov et al., 2004). Рисунок 11 иллюстрирует два примера установленных иерархий гистоновых модификаций, которые, по-видимому, индексируют транскрипцию активного хроматина или, напротив, определяют рисунок гетерохроматиновых доменов.
Эти исследования ставят вопрос о том, существует ли «гистоновый код» или даже «эпигенетический код». Хотя эта теоретическая концепция оказалась весьма стимулирующей, и было показано, что в некоторых своих предсказаниях она правильна, вопрос 6 том, действительно ли существует некий код, остается в значительной степени открытым. Для сравнения скажем, что генетический код оказался крайне полезным ввиду возможности делать предсказания на его основе и благодаря его почти полной универсальности Он использует в основном «алфавит» из четырех оснований в ДНК (т. е. нуклеотидов), образуя в целом инвариантный и почти универсальный язык. В противоположность этому современные данные заставляют считать, что картины гистоновых модификаций значительно варьируют от организма к организму, особенно между низшими и высшими эукариотами, например дрожжами и человеком. Таким образом, даже если гистоновый код и существует, он, вероятно, не универсален. Эта ситуация становится еще более сложной, когда учитывается динамическая природа гистоновых модификаций, варьирующих в пространстве и во времени. Более того, хроматиновая матрица вовлекает в процесс ошеломляющее количество факторов ремоделинга (Vignali et al., 2000; Narlikar et al., 2002; Langst and Becker, 2004; Smith and Peterson, 2005). Однако анализ с использованием иммунопреципитации хроматина (ChIP) в масштабах целого генома (ChIP on chip) начал выявлять неслучайные и в какой-то мере предсказуемые паттерны в нескольких геномах (например, S. pombe, A. thaliana , клетки млекопитающих) — такие как сильные корреляции H3К4me3 с активированными участками промотора (Strahl et al. 1999; Santos-Rosa et al., 2002; Bernstein et al., 2005), a также метилирования H3K9 (Hall et al. 2002; Lippman et al., 2004; Martens et al., 2005) и H3K27 (Litt et al., 2001; Ringrose et al., 2004) с «молчащим» гетерохроматином. Возможно, ограничением гистонового кода является то обстоятельство, что одна модификация не транслируется неизменно в один биологический выходной сигнал. Однако действуя в комбинации или кумулятивным образом, модификации, по-видимому, действительно определяют биологические функции и вносят свой вклад в них (Henikoff, 2005).
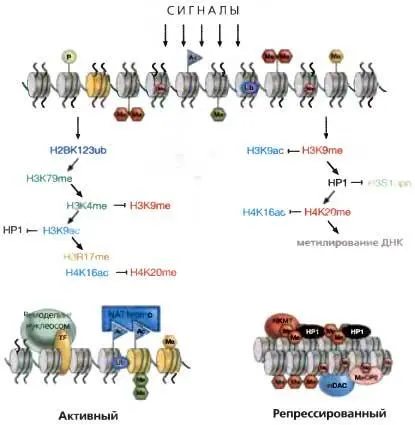
Рис. 3.11.Координированная модификация хроматина
Переход не подвергавшейся воздействию хроматиновой матрицы в активный эухроматин (слева) или образование репрессивного гетерохроматина (справа), включающие ряд скоординированных модификаций хроматина. В случае активации транскрипции это сопровождается действием комплексов ремоделинга нуклеосом и замещением коровых гистонов гистоновыми вариантами (желтый цвет, а именно H3.3)
8. Комплексы, осуществляющие ремоделинг хроматина, и варианты гистонов
Другим важным механизмом, посредством которого индуцируются переходы в хроматиновой матрице, является выработка сигналов [signaling] для рекрутирования комплексов «ремоделинга» хроматина, использующих энергию (гидролиз АТФ) для изменения хроматина и состава нуклеосом нековалентным образом. Нуклеосомы, особенно когда они связаны репрессивными факторами, ассоциированными с хроматином, нередко «навязывают» транскрипционной машине состояние значительного подавления. Отсюда лишь некоторые транскрипционные факторы и регуляторы, специфичные к определенным последовательностям (хотя и не базовая транскрипционная машина), способны получать доступ к сайту (сайтам) своего связывания. Эта проблема доступа решается, хотя бы отчасти, белковыми комплексами, которые мобилизуют нуклеосомы и (или) изменяют нуклеосомную структуру. Ремоделинг хроматина часто функционирует совместно с ферментами, активирующими модификации хроматина, и связанные с ним активности в целом можно разделить на два семейства: семейство SNF2H, или ISWI, и семейство Brahma, или SWI/SNF. Семейство SNF2H/ ISWI мобилизует нуклеосомы вдоль ДНК (Tsukiyama et al., 1995: Varga-Weisz et al., 1997), тогда как Brahma/SWI/ SNF на время изменяет структуру нуклеосомы, экспонируя контакты ДНК: гистон с использованием механизмов, которые только сейчас начинают раскрываться (глава 12).
Кроме того, некоторые из гидролизующих АТФ активностей схожи с «обменными комплексами», которые сами предназначены для замещения обычных коровых гистонов специализированными «вариантными» гистоновыми белками. Эта осуществляемая с затратами АТФ перетасовка в действительности может быть средством замещения существующих модифицированных гистоновых «хвостов» новым, свободным от старого, набором вариантных гистонов (Schwartz and Ahmad, 2005). Альтернативная возможность заключается в том, что рекрутирование таких комплексов ремоделинга хроматина, как SAGA (Spt-Ada-Gcn5-aцeтилтpaнcфepaзa), может быть также усилено пред существующими модификациями гистонов, чтобы обеспечить транскрипционную компетентность промоторов-мишеней (Grant et al., 1997; Hassan et al., 2002).
Читать дальшеИнтервал:
Закладка:



