Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Что представляют собой сигналы, направляющие DNMTs для метилирования определенных участков ДНК? В настоящее время известно, что для того, чтобы «направить» метилирование ДНК de novo, высокоповторяющиеся тандемные последовательности в геноме (например, перицентромерный хроматин) «опираются» на репрессивные метки метилирования по H3К9, как это показано у N. crassa и растений (главы 6 и 9). Рассеянные (interspersed) повторы также могут сигнализировать о метилировании ДНК de novo, как это описано в контексте RIP у N. crassa (Tamaru and Selker, 2001) и сайленсинга ретротранспозонов в зародышевом пути у самцов млекопитающих. Отвечающий за это белок идентифицирован (Dnmt3L) и мог бы функционировать путем сканирования генома для отыскания высоких уровней границ «гомология — гетерология», служащих сигналом о необходимости метилирования ДНК (Bourc’his and Bestor, 2004). У растений сигнал для метилирования ДНК de novo обеспечивают РНК посредством уникального механизма, названного РНК-зависимым метилированием ДНК (RdDM; глава 9). Имеются данные о том, что для глобальных паттернов метилирования ДНК почему-то необходимы комплексы, осуществляющие ремоделинг хроматина и относящиеся к семейству SWI/SNF, как это показано на растениях (Jeddeloh et al., 1999) для белка DDM1 и на млекопитающих с помощью гомолога Lshl (Yan et al. 2003). Наконец, белок EZH2 (относящийся к HKMT-PcG) тоже может участвовать в «направлении» метилирования ДНК в некоторых промоторах у млекопитающих (Vire et al., 2005).
Способ, посредством которого установившееся метилирование ДНК может обусловливать сайленсинг хроматина, выяснен еще не полностью, хотя данные указывают на trans -регуляцию. Белки, связывающиеся с метилированными цитозинами и названные доменными белками, связывающимися с метил-CpG (MBD), могут рассматриваться как основанный на метилировании ДНК эквивалент элементов связывания [binders] (или «считывателей») модифицированных гистоновых мотивов (рис. 3.13). Например, белок, связывающийся с метил цитозином (МеСР2), связывается с метилированными CpG и рекрутирует HDACs, служа связующим звеном для репрессивных гистоновых меток (глава 18) Известно также, что метилирование ДНК нарушает сайты распознавания регуляторов транскрипции (например, СЕСА), участвующих в геномном импринтинге (глава 19).
Наличие метилированной ДНК в импринтированных локусах, обусловливающей сайленсинг либо материнской, либо отцовской аллели у растений и плацентарных млекопитающих, позволяет предположить, что в ходе эволюции эти организмы уникальным образом приспособили этот эпигенетический механизм для стабилизации репрессии генов. Интересно отметить, что у сумчатых метилирования ДНК в импринтированных локусах нет, что указывает на то, что вовлечение этого механизма в импринтинг у млекопитающих является относительно недавним эволюционным событием (главы 17 и 19). Напротив, у таких двукрылых насекомых, как Drosophila , метилирование ДНК как функциональный эпигенетический механизм в основном утрачено (Lyko, 2001).
Высокоповторяющиеся участки генома млекопитающих, обычно метилированные, становятся все более мутагенными, когда они не метилированы, — до такой степени, что вызывают глобальную геномную нестабильность (Chen et al., 1998). В результате возникают хромосомные аномалии, являющиеся главной причиной многих болезней и прогрессии рака (см. раздел 15). Это подчеркивает решающую роль метилирования ДНК в обеспечении целостности генома. С другой стороны, отдельные метилированные остатки цитозинов весьма предрасположены к спонтанному мутированию. Таким образом, со временем в результате реакции дезаминирования возникают транзиции С-Т (рис. 3.13), но полагают, что эта характеристика благоприятна для защиты хозяйского генома, потому что она постоянно дезактивирует такие паразитические нуклеотидные последовательности ДНК, как транспозоны. В ином контексте эта же самая химическая реакция активно катализируется дезаминазой, индуцируемой активацией, или AID (activation-induced deaminase). Экспрессия этого энзима в В- и Т-клетках вызывает «соматические гипермутации» в иммуноглобулиновом локусе (lg). Это важный механизм расширения репертуара рецепторов антигенов и, отсюда, усиления иммунитета у млекопитающих (Petersen-Mahrt, 2005; Глава 21). Экспрессия AID, наблюдаемая в раннем развитии млекопитающих, привела к предположению, что она может обеспечивать альтернативный путь к деметилированию ДНК, хотя это происходило бы на фоне риска повышенной частоты точечных мутаций.
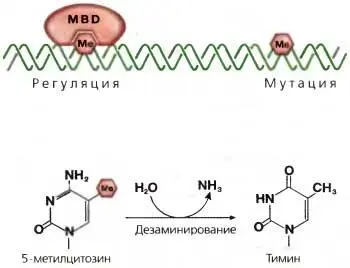
Рис. 3.13.Метилирование и дезаминирование ДНК
Метилированные (Ме) цитозиновые нуклеотиды могут быть связаны с метилДНК-связывающимися белками (MBD) Несвязанный 5-метилцитозин склонен спонтанно мутировать в результате дезаминирования (реакция, изображенная в нижней части рисунка); в результате в нуклеотидной последовательности ДНК происходит траH3иция 5-метил-СрС в ТрА
Метилирование ДНК и метилирование гистонов являются известными механизмами эпигенетической регуляции генома Некодирующие РНК, как описано в следующем разделе, являются важными первичными триггерами для индукции «молчащего» хроматина. Известно также, что молекулы РНК могут быть интенсивно метилированы по сахарному или нуклеозидному скелету. Более того, показано, что метилирование малых некодирующих РНК на З’-конце стабилизирует эти молекулы (Yu et al., 2005). Любопытно, что Dnmt2 недавно была идентифицирована как метилтрансфераза тРНК (Goll et al., 2006). Поэтому похоже, что РНК-метилтрансферазы, подобные DNMTs и HKMTs, могут существовать как «считыватели» эпигенетической информации, хотя прямых данных, подтверждающих это, нет. Однако метилирование РНК, по-видимому, «чувствуется» определенными Toll-подобными рецепторами (трансмембранными рецепторами, которые распознают молекулярные мотивы обычных патогенов), чтобы опосредовать врожденный иммунитет (Ishii and Akira, 2005), что говорит в пользу такой гипотезы. Возникает интересная возможность, что метилирование РНК может все же быть эпигенетической модуляцией на основе третьей формы метилирования.
10. РНКи и сайленсинг генов, направляемый РНК
Сведения о том, что конститутивный гетерохроматин в центромерах и теломерах играет инструктивную роль в обеспечении целостности генома, способствовали сдвигу парадигмы во взглядах на повторяющуюся некодирующую «мусорную» ДНК. Возможно ли, что эти повторяющиеся последовательности служат каким-то «незряшным» целям, которые еще только начинают проясняться? Не может ли даже быть так, что эти нуклеотидные последовательности ДНК не являются полностью «молчащими»?
Такая возможность вытекает из фундаментальной серии открытий, связывающих РНКи с образованием «молчащего» хроматина (гетерохроматина). РНКи является защитным механизмом организма-хозяина, который разбивает dsRNA на мелкие молекулы РНК (известные как короткие интерферирующие РНК или siRNA). Этот процесс в конечном счете приводит к деградации РНК или к использованию этих малых РНК для подавления трансляции, известного как посттранскрипционный сайленсинг (PTGS). Открытый позже механизм транскрипционного сайленсинга генов (TGS), ведущего к образованию гетерохроматина, был обнаружен в результате конвергенции независимых линий исследования хроматина и механизма РНК-интерференции. С одной стороны, многое известно о репрессивном метилировании ДНК (у грибов, растений и млекопитающих), модификациях хроматина (например, H3К9me3) и ассоциированных с хроматином факторах (НР1), характерных для доменов гетерохроматина. С другой стороны, исследователи преуспели в идентификации факторов механизма РНКи (например, Dicer, Argonaute, РНК-зависимой РНК-полимеразы, или RdRP). Наиболее убедительный прогресс, связавший вместе эти вроде бы разные области, был достигнут в элегантных исследованиях на S. pombe, в которых мутации любого компонента механизма РНКи приводили к дефектам в расхождении хромосом (Hall et al., 2002; Reinhart and Bartel, 2002; Volpe et al., 2002). Это было вызвано неспособностью стабилизировать центромерный гетерохроматин и подчеркивало, по-видимому, широко распространенную роль механизмов, опосредованных РНКи, в образовании доменов «молчащего» гетерохроматина. Это также высветило значение гетерохроматина помимо транскрипционного сайленсинга генов — его роль в поддержании целостности генома и, отсюда, жизнеспособности, как показывает необходимость центромерного гетерохроматина для процесса расхождения хромосом. Появляющиеся данные позволяют также предполагать, что siRNAs требуются для определения других специализированных районов функционального гетерохроматина, таких как теломеры.
Читать дальшеИнтервал:
Закладка:



