Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Такие cis -активные элементы в ДНК могли бы функционировать посредством сиквенс-специфичного связывания белка, способного включать формирование гетерохроматина Были идентифицированы белки, специфически связывающиеся с некоторыми из сателлитных ДНК (например, D1 — Aulner et al., 2002). Вывод о важности этих взаимодействий был сделан, исходя из влияния специфичных для сателлитной ДНК веществ, связывающихся с ДНК и способных подавлять PEV (Janssen et al., 2000). Однако данные, полученные на дрожжах и растениях (Elgin and Grewal, 2003; Matzke and Birchler, 2005; см. Главы 8 и 9), позволяют предложить другую модель, а именно, что некий основанный на RNAi механизм мог бы использоваться для «нацеливания» формирования гетерохроматина на повторяющиеся элементы. Работы из нескольких лабораторий показали, что система RNAi у Drosophila имеется и играет важную роль в регуляции в ходе развития посредством посттранскрипционного сайленсинга генов (PTGS). D. melanogaster обладает двумя генами, кодирующими белки DICER, и многочисленными генами (aubergine, AGO1, AGO2, spindle [известный также как homeless ], vasa intronic gene [ VIG ], armitage, Fmr1 ), кодирующими компоненты или белки, необходимые для сборки индуцируемого РНК сайленсирующего комплекса (RISC) (Sontheimer, 2005). Участие этой системы предположили в PTGS повторяющихся последовательностей, особенно тандемно повторяющихся генов Stellate , нескольких ретротранспозонов и трансгенов Alcohol dehy-rogenase (Adh) и в транскрипционном сайленсинге (TGS) трансгенов Adh (Aravin et al., 2001; Pal-Bhadra et al., 2002). Пал-Бхадра с соавторами (Pal-Bhadra et al., 2004) обнаружили при прямом тестировании, что мутации в piwi (член семейства домена PAZ) и homeless (DEAD-боксовая геликаза) подавляют PEV, связанный с тандемными порядками гена white, и что мутации в piwi, aubergine и homeless подавляют сайленсинг трансгена white P [ hsp70-w ] в перицентромерном гетерохроматине или четвертой хромосоме. Эта супрессия PEV была связана со значительным снижением уровня метилирования H3K9. Связанные с повторами малые интерферирующие РНК (rasiRNAs) были идентифицированы с 40 % известных мобильных элементов (включая 1360) и других повторяющихся последовательностей (Aravin et al., 2003).
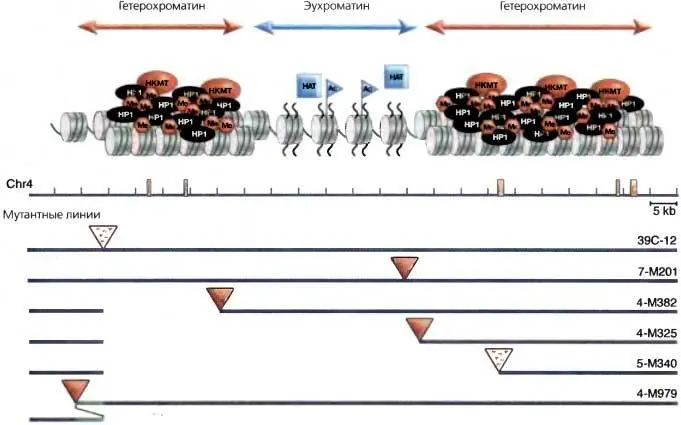
Рис. 5.7.Возможная модель гетерохроматинового «нацеливания»
dsRNA с повторяющихся последовательностей процессируется с помощью RISC и генерирует гипотетический «комплекс нацеливания», который управляет либо модификацией гистонов, либо ассоциацией с НР1 как начальным этапом в сборке гетерохроматина в сайте, идентифицированном малой ssRNA. Данные, полученные на четвертой хромосоме, позволяют предположить, что фрагменты ДНК-транспозона 1360 ( оранжевые полосы ) являются мишенью для формирования гетерохроматина: локальные делеции или дупликации, которые сдвигают положение репортера R-элемента ( треугольник ) в сторону от элемента 1360, ведут к утрате сайленсинга ( красный треугольник обозначает красный глаз ), тогда как близость к 1360 приводит к сайленсингу ( треугольник с точками обозначает мозаичный глаз ) (на основе данных Sun et al., 2004)
В совокупности эти обсуждавшиеся выше результаты заставляют предполагать, что формирование гетерохроматина может зависеть как от ядерной локализации (возможно, создающих богатый пул требующихся белков), так и от специфического «нацеливания» на основе RNAi-распознавания и процессинга двунитевой РНК с повторяющихся элементов, в особенности некоторых из ДНК-транспозонов. Такое «нацеливание» посредством RISC могло бы привлечь либо метилтрансферазу гистона H3, либо комплекс, включающий НР1 (либо и то, и другое) к сайту для включения процесса сборки, обсуждавшегося в разделе 4.
7. Не весь гетерохроматин идентичен
Хотя в общих терминах гетерохроматин был описан выше, ясно, что гетерохроматиновые домены варьируют в деталях. Все гетерохроматиновые домены характеризуются повторяющейся ДНК (см. выше), но это может варьировать от тандемных последовательностей коротких повторов (сателлитная ДНК), обнаруживаемых в блоках в центромерных районах, до высокой плотности перемежающихся повторяющихся последовательностей, что наблюдается в четвертой хромосоме. В то время как все гетерохроматиновые районы, по-видимому, ассоциированы с НР1 и H3K9me2, ясно, что связанные с этим белковые комплексы должны отличаться в иных отношениях. Исследование влияния 70 разных модификаторов на разные гены, демонстрирующие мозаицизм (в том числе w т4, bw D, репортеры R-элемента в перицентромерном гетерохроматине или в порядке [array] TAS), показало, что на фоне существенного перекрывания в мишенях модификаторов имеет место удивительная сложность. С помощью использованного набора тестов эти модификаторы разделили на семь разных групп с точки зрения их способности влиять на сайленсинг в данном компартменте (Donaldson et al., 2002). Интересно, что единственным модификатором в этой группе, влияющим на сайленсинг в порядке TAS, была новая аллель Su(var)3-9.
Эти различия несомненно отражают изменения в локальной биохимии или в энзимах, используемых для этого. Например, цитологические результаты показывают, что в то время, как H3K9me2 сильно концентрируется вдоль по длине четвертой хромосомы, ответственный за это энзим — это не SU(VAR)3-9 (Schotta et al., 2002; К.A. Haynes et al., неопубликовано). Даже в пределах перицентромерного гетерохроматина следует ожидать мозаицизм, принимая во внимание различия в составляющих его основу блоках ДНК, которые варьируют от сателлитной ДНК до кластеров перемежающихся повторов (Le et al., 1995), которые могли бы использовать разную смесь гетерохроматиновых белков. Последствия были видны в исследованиях, где изучали влияние разных блоков перицентромерного гетерохроматина на экспрессию с репортера, и можно было наблюдать, что степень выраженности [severity] фенотипа зависит не просто от количества гетерохроматина в cis -конфигурации, но варьирует в зависимости от локального гетерохроматинового окружения (Howe et al., 1995). Связанные с гетерохроматином белки, которые могут играть роль в специфических субдоменах, включают AT-hook белок D1, преимущественно связанный с сателлитом III с плотностью 1,688 г/см 3(Aulner et al., 2002), и DDP1, мульти-КН-доменный белок, гомологичный вигилину, который связывается с богатой пиримидинами С-нитью додекасателлита (Cortes and Azonn, 2000).
8. PEV, формирование гетерохроматина и сайленсинг генов у различных организмов
Явление мозаицизма, обусловленного эффектом положения (PEV), первоначально было установлено на Drosophla просто потому, что это был один из первых организмов, на которых для получения мутаций было использовано Х-излучение. Х-излучение с гораздо большей вероятностью, чем другие обычно используемые мутагены, индуцирует хромосомные перестройки, результатом которых может быть PEV. Сходные мутации были изолированы у мышей, где пятнистая окраска меха указывает на PEV. Генетический анализ выявил вставку аутосомного района, несущего аллели дикого типа генов окраски меха, в Х-хромосому (Cattanach, 1961; Russel and Bangham, 1961). Мозаичность наблюдается только у самок, несущих эту вставку в сочетании с гомозиготной мутацией в исходных генах окраски меха. У этих самок аллель дикого типа становится инактивированной вследствие Х-инактивации путем гетерохроматинизации (глава 17). У растений единственный несомненный случай PEV был описан у Oenothera blandina (Catcheside, 1939). В этих случаях, как и у Drosophila, PEV-сайленсинг эухроматиновых генов связан с перемещением этих генов в новое гетерохроматиновое окружение.
Читать дальшеИнтервал:
Закладка:



