Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
PEV отражает изменение в экспрессии гена вследствие генетической перестройки, а именно утрату экспрессии репортерного гена в некоторых клетках, в которых он в норме активен. Для объяснения PEV предложено несколько разных моделей; не все из них являются взаимоисключающими. Одна из возможностей, рассмотренная исходно, — это случайная потеря гена, возможно вследствие поздней репликации (Karpen and Spradling, 1990). Анализ с помощью количественного Саузерн-блоттинга показал, что это объяснение в общем виде неприменимо; гены, обнаруживающие мозаицизм, обычно полностью реплицируются в диплоидной ткани (Wallrath et al., 1996). Другие модели делают акцент на ассоциации гена, обнаруживающего мозаицизм, с гетерохроматиновым компонентом ядра и (или) на распространении гетерохроматиновой структуры с гетерохроматина, оказавшегося по соседству. Эта модель распространения, базирующаяся на многочисленных генетических и цитологических данных, объясняет сайленсинг как следствие распространения гетерохроматиновой упаковки через точку разрыва в эухроматиновые в норме районы. В нормальных хромосомах эухроматиновые и гетерохроматиновые районы, по-видимому, изолированы [insulated] друг от друга специфическими последовательностями или буферными зонами. Поскольку эти «изолирующие последовательности» [«insulating sequences»] (которые никогда не были полностью определены у Drosophila) отсутствуют в местах соединения эухроматина и гетерохроматина при PEV-перестройках (см. рис. 5.1), гетерохроматинизация эухроматиновых последовательностей индуцируется с переменным успехом. Эту гетерохроматинизацию можно видеть на цитологическом уровне в политенных хромосомах как сдвиг от бэндированной к аморфной структуре в основании хромосомных плечей (Hartmann-Goldstein, 1967); степень этого изменения можно модифицировать мутациями Su(var) и E(var) (Reuter et al., 1982).
Инактивацию эухроматиновых генов на расстоянии вдоль по длине хромосомы можно продемонстрировать генетически (Demerec and Slizynska, 1937). Затронутые районы становятся связанными с НР1 (Belyaeva et al., 1993) и обнаруживают H3K9me2, типичную метку гетерохроматина Drosophila (Ebert et al., 2004). Поскольку модель распространения постулирует конкуренцию между упаковкой в эухроматин и упаковкой в гетерохроматин, гены-модификаторы PEV могли бы кодировать функции контроля либо формирования гетерохроматина, либо формирования эухроматина. Обнаружение зависимых от дозы модификаторов, как обсуждается выше, свидетельствует в пользу такой модели (Locke et al., 1988; Henikoff, 1996). И действительно, недавно были идентифицированы мутации Su(var), контролирующие баланс между эухроматином и гетерохроматином (Ebert et al., 2004). PEV-перестройки позволили нам визуализировать и изучить случаи, где гетерохроматиновая упаковка распространяется на фланкирующий эухроматиновый домен. Этот эффект распространения несомненно зависит от ряда молекулярных реакций в эухроматиновых районах. Известны несколько модификаций гистонов, которые являются взаимоисключающими и которые определяют эти альтернативные хромосомные состояния. Ацетилирование H3K9, метилирование H3K4 и фосфорилирование H3S10 — типичные метки активного эухроматина, тогда как метилирование H3K9, H3K27 и H4K20 является специфической меткой «молчащих» районов. Следовательно, гетерохроматинизация эухроматиновых районов требует специфических реакций деацетилирования, деметилирования и дефосфорилирования в эухроматине, как это изображено на рис. 5.6. Этот переход исходно зависит от деацетилирования H3K9 ферментом HDAC1 Мутации в гене rpd3 , кодирующем специфическую для H3K9 деацетилазу гистонов HDAC1, являются сильными супрессорами PEV(Mottus et al., 2000), выступая антагонистами эффекта SU(VAR)3-9 в сайленсинге генов (Czermin et al., 2001). Как было показано, HDAC1 in vivo ассоциирован с комплексом SU(VAR)3-9/HPl; эти два энзима работают кооперативно, метилируя предацетилированные гистоны.
Недавно наблюдали, что распространение гетерохроматина в эухроматин полностью блокируется у мутантов Su(var)3-1 (Ebert et al., 2004). Мутации Su(var)3-1 являются мутациями сдвига рамки считывания в гене, кодирующем киназу JIL1; они приводят к экспрессии усеченного белка JIL1, не имеющего карбокситерминального района. Белок JIL1 содержит два киназных домена и катализирует фосфорилирование H3S10 в эухроматине. Мутации JIL1 Su(var)3~ 1 не влияют на фосфорилирование H3S10, но, вероятно, нарушают дефосфорилирование H3S10, эффективно подавляя метилирование H3K9. Это заставляет предположить участие фосфатазы. Неизвестно, участвует ли энзим РР1 (который был идентифицирован с помощью мутаций Su(var)3-5) (Baksa et al., 1993) непосредственно в этой реакции. Деметилирование H3K4 является, по-видимому, еще одной предпосылкой для гетерохроматинизации эухроматиновых районов. Недавние работы показали, что аминооксидаза LSD1 функционирует у млекопитающих как деметил аза H3K4 (Shi et al., 2005). Предполагаемый гомолог LSD1 у Drosophila, SU(VAR)3-3, облегчает распространение гетерохроматина в эухроматиновые районы во всех испытанных PEV-перестройках (S. Lein et al., неопубликовано). В соответствии с этим в нуль-клетках по Su(var)3-3, у которых нет LSD1, приобретение метилирования H3K9 в эухроматине, фланкирующем точку разрыва, отменяется, хотя конститутивно гетерохроматиновые районы не затрагиваются. Эти открытия показывают, что для удаления специфических для эухроматина меток модификаций гистонов необходима координированная функция нескольких энзимов, прежде чем может иметь место переход к гетерохроматиновой упаковке (см. рис. 5.6). Вполне вероятно, будет обнаружено, что эти необходимые энзимы образуют комплексы с SU(VAR)3-9/HPl, как это было уже показано для HDAC1.
6. Как формирование гетерохроматина «нацеливается» у Drosophila ?
Хотя мы многое узнали о механистических аспектах и биохимии структуры гетерохроматина, как это описано выше, остается открытым вопрос о том, каким образом формирование гетерохроматина «нацеливается» на избранные районы генома в его нормальной конфигурации. Все гетерохроматиновые домены обладают некоторыми общими чертами, и две такие особенности признаны существенными входными сигналами [inputs] для сборки гетерохроматина на данной нуклеотидной последовательности ДНК: положение локуса относительно пространственно обособленных [distinct] субдоменов гетерохроматина в ядре и присутствие повторяющейся ДНК
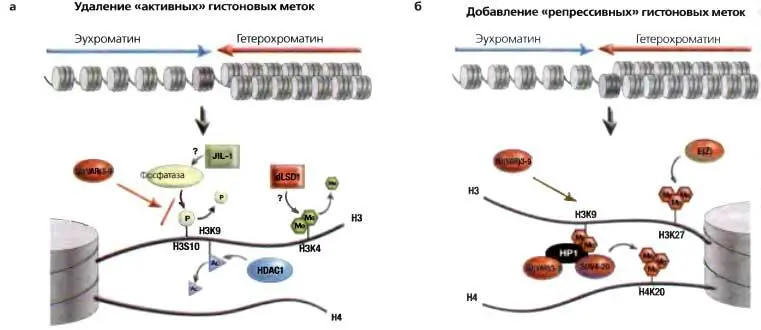
Рис. 5.6.Переход из эухроматинового состояния в гетерохроматиновое состояние требует ряда изменений в модификациях гистонов
(а) Активные гены маркированы H3K4me2 и me3; эта метка, если присутствует, предположительно должна быть удалена LSD1 (у Drosophila пока еще не охарактеризован). В эухроматине НЗК9 в норме апетилирован; эта метка должна быть удалена деацетилазой гистонов, HDAC1. Фосфорилирование H3S10 может интерферировать с метилированием НЗК9; дефосфорилирование, по-видимому, происходит с участием фосфатазы, «нацеливаемой» взаимодействием с карбоксильным концом киназы JIL1 Эти переходы создают обстановку для приобретения модификаций, связанных с сайленсингом (показано на б ), в том числе метилирования НЗК9 белком SU(VAR)3-9, связывания НР1 и последующего метилирования Н4К20 белком SUV4-20, ферментом, рекрутируемым НР1. Может также происходить метилирование НЗК27 энзимом E(Z)
Читать дальшеИнтервал:
Закладка:



