Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
В перестройке w m4 ген white в результате инверсии оказывается в непосредственной близости с гетерохроматиновым материалом Х-хромосомы, локализованным на дистальной границе ядрышкового организатора (Cooper, 1959). Этот район содержит тандемные порядки мобильных элементов типа R1; гетерохроматиновая точка разрыва In(1)w m4 находится внутри единицы повтора R1 (A. Ebert and G. Reuter, неопубликовано). После воздействия Х-лучами или EMS (этанметилсульфонат, химический мутаген) были изолированы ревертанты w m4, имеющие фенотип w + (Tartof et al., 1984; Reuter et al., 1985). Анализ более 50 ревертантных хромосом w + (все они обнаружили реинверсию или транслокацию гена white в соседство с эухроматином) позволил предположить, что гетерохроматиновый материал, фланкирующий непосредственно точку разрыва, вызывает инактивацию гена white у w m4. Большинство ревертантов снова обнаруживают белую пятнистость, если вводятся сильные мутации E(var), что заставляет предполагать, что некоторые гетерохроматиновые последовательности после перемещения остаются ассоциированными с геном white. Эти исследования позволяют считать повторяющуюся ДНК (в данном случае повторы R1) мишенью для формирования гетерохроматина.
Большинство известных мутаций-модификаторов PEV были изолированы с использованием сенсибилизированного генетического фона. Для изоляции доминантных супрессорных мутаций тест-генотип содержит доминантный энхансер, тогда как в схемах для изоляции энхансерных мутаций используется доминантный супрессор (Dorn et al., 1993b). Если тест-генотип содержит энхансер мозаичности, все мухи w т4 имеют белые глаза, и исключения, имеющие пятнистые или красные глаза, указывают на вновь индуцированные мутации Su(var). Соответственно, в тест-линии с доминантным супрессором все мухи w т4 имеют красные глаза, а исключительные мухи с мозаичным фенотипом указывают на вновь индуцированные мутации E(var). Эти сенсибилизированные генетические схемы благоприятствуют изоляции сильно доминантных мутаций Su(var) и E(var), которые оказались очень полезными для детального генетического анализа.
С использованием этого подхода в разных опытах по скринингу просмотрели более миллиона мух, и были изолированы более 140 мутаций Su(var) и 230 мутаций
E(var) (Schotta et al., 2003). Мутации были индуцированы EMS, Х-лучами или ремобилизацией элементов Р. Еще один набор мутаций Su(var) был изолирован в ходе прямого скрининга с w m4 (Sinclair et al., 1983). Скрининг с хромосомой Df(l'J), демонстрирующей сильный мозаицизм по гену yellow (маркер, желтая окраска тела), привел к изоляции 70 мутаций-модификаторов PEV (Donaldson et al., 2002). Кроме того, скрининг на доминантные модификаторы экспрессии транспозонного репортерного гена позволил идентифицировать несколько мутаций с эффектом Su(var) (Birchler et al., 1994). Известно, что выборка критических регуляторных генов даун — регулируется генами группы Polycomb (PcG) и ап — регулируется генами группы trithorax (trxG). В прямых тестах относительно немногие мутации в генах PcG модифицируют PEV (например, Sinclair et al., 1998). Напротив, многие мутации в генах trxG являются энхансерами PEV (Dorn et al., 1993а; Farkas et al., 1994).
В совокупности этот скрининг позволил идентифицировать большое число доминантных мутаций Su(var) и E(var). По данным выполненного к настоящему времени генетического анализа общее число мутаций Su(var) и E(var) можно оценить как приблизительно 150. Большое число генов Su(var) и E(var) с почти идентичными фенотипическими эффектами иногда приводит к несоответствиям в генетической номенклатуре. Чаще всего символы генов Su(var) и E(var) комбинируются с цифрами, указывающими хромосому, в которой локализована данная мутация, с номером гена и номером аллели Так, Su(var)3-9 17 символизирует аллель 17 девятого гена Su(var), идентифицированного в третьей хромосоме. В настоящее время были тщательно картированы лишь около 30 соответствующих генов и идентифицированы соответствующие аллели (табл. 5.1). Зависящие от дозы эффекты наблюдались для примерно одной трети идентифицированных генов, с использованием серии перекрывающихся нехваток и дупликаций. В этих случаях уменьшение количества генных продуктов из-за утери одной копии гена неизменно приводит к модификации мозаичного фенотипа. Делеции этих локусов Su(var) или E(var) супрессируют или, соответственно, усиливают сайленсинг генов. Исследования с использованием дупликаций позволили идентифицировать новые локусы-модификаторы, которые обнаруживают противоположное (антиподное) влияние на PEV, если путем дупликации или с помощью инсерции трансгена вводится лишняя копия данного гена. Общее число генов-модификаторов PEV, обнаруживающих зависящие от дозы эффекты, оценивается примерно в 15—20 (Schotta et al., 2003).
Если утрата одной копии гена приводит к супрессии PEV, а присутствие трех копий гена ведет к усилению PEV, это позволяет предполагать, что для установления гетерохроматина требуется, в стоихиометрических количествах, продукт, кодируемый этим геном, с сопутствующим сайленсингом гена (рис. 5.2). Были детально охарактеризованы три таких локуса, Su(var)2-5 (кодирующий НР1), Su(var)3-7 (кодирующий белок типа «цинковых пальцев») и Su(var)3-9 (кодирующий метилтрансферазу лизина гистонов). Su(var)2-5 был клонирован путем скрининга библиотеки экспрессии кДНК с помощью моноклонального антитела, распознающего гетерохроматин (James and Elgin, 1986). Кодируемый белок, ассоциированный с гетерохроматином, был впоследствии обозначен НР1 (белок гетерохроматина 1). Гибридизация in situ с использованием выделенной клонированной ДНК позволила идентифицировать ген в районе 28—29 политенных хромосом,
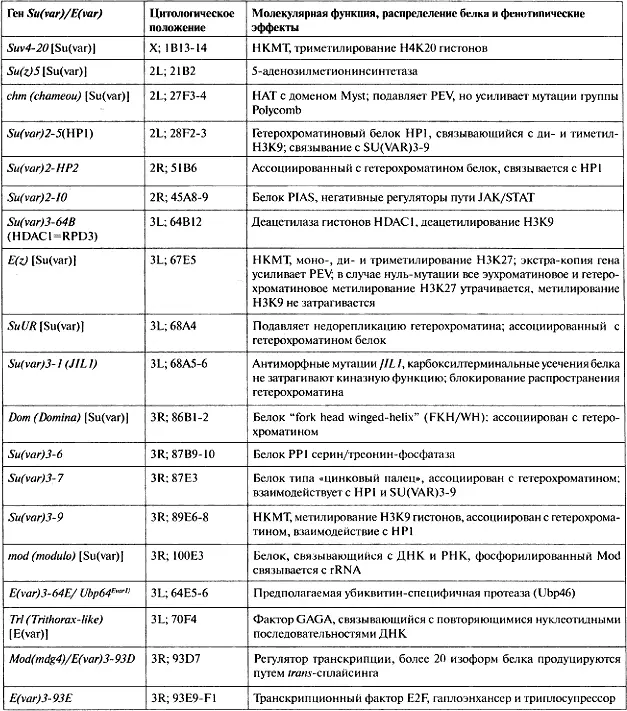
См. оригинальное цитирование во Flybase.
где ранее был картирован Su(var)2-5- Анализ нуклеотидной последовательности ДНК мутантных аллелей подтвердил, что локус Su(var)2-5 в положении 28F1-2 на хромосоме кодирует HP 1 (Eissenberg et al., 1990), HP1 содержит два консервативных домена, аминотерминальный хромодомен и карбокситерминальный домен «chromoshadow» (Paro and Hogness, 1991), и взаимодействует с несколькими другими хромосомными белками. Su(var)3- 7 был сперва цитогенетически картирован (с помощью серии перекрывающихся делеций и дупликаций) в районе 87Е1-4 в третьей хромосоме. Этот район был проанализирован на уровне ДНК как часть первой «прогулки по хромосоме» [chromosomal walk], выполненной у Drosophila (Bender et al., 1983). С помощью серии перекрывающихся геномных клонов Su(var)3-7 был определен в пределах фрагмента ДНК величиной 7.8 т.о., который обладал трипло-энхансерным влиянием на репортер, обнаруживающий мозаичность (Reuter et al., 1990). Su(var)3-7 кодирует белок с семью регулярно расположенными [regularly spaced] «цинковыми пальцами» — доменами, которые, как было показано, функционируют в связывании с ДНК (Cleard and Spierer, 2001). Su(var)3-9 был клонирован с помощью таггирования транспозоном P-элементом (Tschiersch et al., 1994). Ген Su(var)3-9 у Drosophila (и у всех других голометаболических насекомых, изученных к настоящему времени) образует бицистронную единицу с геном, кодирующим eIF2y (Krauss and Reuter, 2000). Поскольку транскрипционная единица Su(var)3-9 не имеет интронов, вполне вероятно, что Su(var)3-9 был вставлен в интрон гена eIF2y путем ретротранспозиции. Белок SU(VAR)3-9 содержит хромодомен в своем аминотерминальном участке и домен SET (идентифицированный впервые в белках SU(VAR)3-9, ENHANCER OF ZESTE [E(Z)] и TRITHORAX) (Jones and Gelbart, 1993; Tschiersch et al., 1994) на своем карбоксильном конце. Этот белок является метилтрансферазой гистонов, специфически модифицирующей гистон H3 по лизину 9.
Читать дальшеИнтервал:
Закладка:



