Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Были описаны семь разных мутантных аллелей Su(var)2-5 , в том числе миссенс-мутации в хромодомене, преждевременные стоп-кодоны и ошибки сплайсинга (Eisenberg et al., 1992). Мутации Su(var)3-7 были получены с помощью гомологичной рекомбинации (Seum et al., 2002); дополнительные аллели были выявлены как супрессоры сайленсинга, зависимого от P-элемента (Bush-еу and Locke, 2004). Сорок мутантных аллелей Su(var)3-9 были выявлены и определены на молекулярном уровне (Ebert et al., 2004). Иммуноцитологический анализ с использованием специфических антител или химерных белков слияния, экспрессируемых трансгеном [transgene-expressed fusion proteins], показал, что все три белка преимущественно ассоциированы с гетерохроматином (см. ниже и рис. 5.4) (James et al., 1989; Cleard et al., 1997; Schotta et al., 2002). Сильная колокализация особенно очевидна для НР1 и SU(VAR)3-9. Эти белки также связываются с теломерами и в ряде эухроматиновых сайтов (Fanti et al., 1998; Schotta et al., 2002).
Несколько инсерций P- элемента, несущих репортерный ген w + в теломерные районы, обнаруживают белую пятнистость. Этот феномен называется теломерным эффектом положения (ТРЕ). Упаковка, подобная гетерохроматину, наблюдается в ассоциированных с теломерами сателлитных последовательностях (TAS), кластерах повторяющейся ДНК, расположенных проксимально по отношению к ретровирусным элементам НеТ-А и TART, составляющим теломеры Drosophila (Cryderman et al., 1999a). В целом, не было обнаружено модификации ТРЕ мутациями в известных генах-модификаторах, хотя НР1 важен для целостности теломер. В клетках с недостаточностью по этому белку хромосомы часто сливаются в области своих теломер (Fanti et al., 1998). У Drosophila в ходе недавнего скрининга не было обнаружено никаких /гая5-действуюших доминантных модификаторов ТРЕ (Mason et al., 2004), что заставляет предполагать, что эти участки сайленсированы двумя (или несколькими) независимыми механизмами.
3. Иммунофлуоресцентное окрашивание политенных хромосом позволило идентифицировать белки, специфически ассоциированные с гетерохроматином
Одним из преимуществ работы с Drosophila является возможность изучения политенных хромосом, дающая визуальную «дорожную карту» генома. На стадии личинки хромосомы во многих терминально дифференцированных клетках реплицируются, но не проходят через митоз; нити хроматина остаются спаренными, в состоянии совершенного синапсиса, и все копии выравнены друг с другом. Наиболее крайний случай имеет место в слюнных железах, где эухроматиновые плечи хромосом претерпели 10 раундов репликации, дав в результате около 1000 копий. Однако репликация не является однородной; многие повторяющиеся последовательности недореплицируются, а сателлитные последовательности ДНК не реплицируются вовсе Все хромосомные плечи сливаются в общем хромоцентре. Таким образом, у D. melanogaster наблюдаются пять длинных плечей (X, второе левое [2L], второе правое [2R], третье левое [3L], третье правое [3R]) и короткое плечо четвертой хромосомы, выступающее из конденсированного хромоцентра, состоящего из перицентромерного гетерохроматина (рис. 5.4а) (обзор см. Ashburner et al., 2005)
Хотя генетический анализ позволил идентифицировать многие локусы, необходимые для формирования гетерохроматина, сам по себе он не позволяет определить, играет ли продукт данного локуса прямую или же косвенную роль. Специфическая ассоциация белка с гетерохроматином первоначально наблюдалась при скрининге моноклональных антител (полученных с использованием прочно-связывающихся ядерных белков, где анализировали паттерны распределения на политенных хромосомах. Антитела, специфичные к белку 22 кДа, впоследствии обозначенному как НР1, давали иммунофлуоресцентное «окрашивание» перицентромерного гетерохроматина, теломер и бэндированной [banded] части маленькой четвертой хромосомы, т.е. известных сайтов гетерохроматина (рис. 5.4а) (James and Elgin, 1986). Последующий анализ (описанный выше) показал, что белок НР1 кодируется Su(var)2-5 , известным супрессором PEV (Eissenberg et al., 1990). Изучение хромосомной локализации специфическими антителами с использованием либо митотических хромосом (Fanti and Pimpinelli, 2004), либо политенных хромосом (обеспечивающих большее разрешение, но недостаточных по центромерному гетерохроматину) (Silver and Elgin 1976) остается наилучшим способом демонстрации того, что продукт локуса Su(var) кодирует хромосомный белок. Были идентифицированы приблизительно 10 таких специфичных для гетерохроматина белков; если есть мутации в генах, кодирующих эти белки, часто наблюдают доминантную супрессию PEV (см. табл. 5.1) (Ashburner et al., 2005b). Эти белки, в том числе недавно идентифицированный НР2 (рис. 5.4а) (Shaffer et al., 2002), являются кандидатами на роль структурных компонентов гетерохроматина.
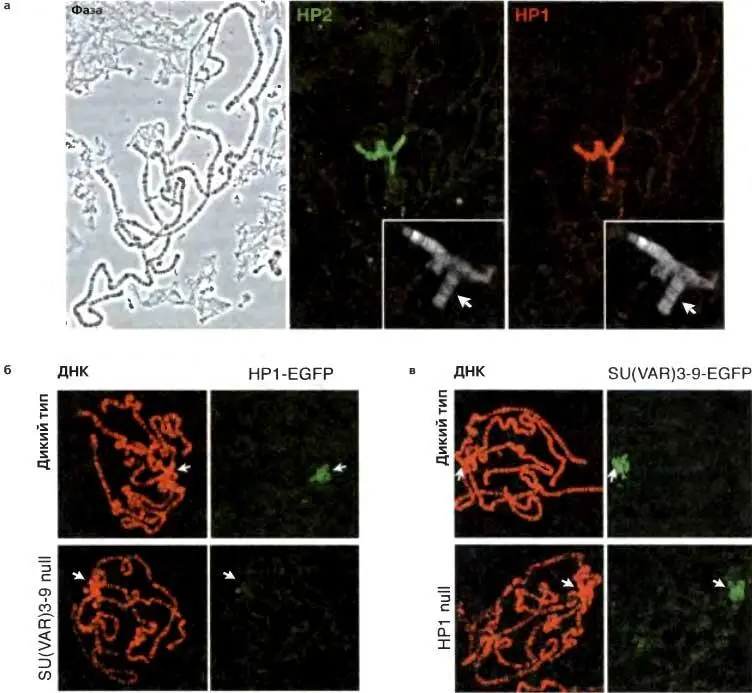
Рис. 5.4.Иммунофлуоресцентное окрашивание политенныххромосом позволяет идентифицировать белки, преимущественно ассоциированные с гетерохроматином
(а) Политенные хромосомы, на фиксированных и давленых препаратах слюнных желез (микроскопия с фазовым контрастом, слева) «окрашены» с помощью инкубирования сначала с антителами, специфичными к данному хромосомному белку, а затем со вторичным антителом, конъюгированным с флуоресцентной меткой. НР1 ( справа ) и НР2 ( в центре ) имеют сходный рисунок распределения, демонстрирующий бросающуюся в глаза связь с перицентромерным гетерохроматином, маленькой четвертой хромосомой ( врезка, стрелка ) и небольшой группой сайтов в эухроматиновых плечах. Обратите внимание на то, что на эффективность любого антитела может влиять выбор протокола фиксации (см. Stephens et al. 2003). (б, в) Ассоциация НР1 и SU(VAR)3-9 с перицентромерным гетерохроматином является взаимозависимой. Мутации в Su(var)3-9 приводят к потере НР1 из перицентромерного гетерохроматина (но не четвертой хромосомы; см. текст) (б), тогда как мутации в Su(var)2-5 приводят к делокализации SU(VAR)3-9 (в) (из: Shatter et al. 2002, с изменениями)
4. Модификация гистонов играет ключевую роль в сайленсинге гетерохроматина
Анализ SU(VAR)3-9 позволил идентифицировать ключевую функцию, необходимую для сайленсинга гетерохроматиновых генов (Tschiersch et al., 1994). Этот белок содержит домен SET, функционирующий как фермент в метилировании H3K9 гистонов. То, что этот белок является метилтрансферазой лизина гистонов (HKMT), «нацеленной» на H3K9, было впервые показано в ходе изучения гомологичного белка человека, SUV39H1 (Rea et al., 2000). У Drosophila SU(VAR)3-9 является главной, но не единственной HKMT H3K9 (Schotta et al., 2002; Ebert et al., 2004). SU(VAR)3-9 контролирует диметилирование H3K9в основной массе перицентромерного гетерохроматина, но не в четвертой хромосоме, теломерах или эухроматиновых сайтах. Триметилирование H3K9, которое у Drosophila наблюдется, главным образом, во внутреннем хромоцентре, также контролируется SU(VAR)3-9. Диметилирование этого внутреннего района не зависит от SU(VAR)3-9, как и монометилирование H3K9 в перицентромерном гетерохроматине (Ebert et al., 2004). HKMTs, ответственные за эти модификации, все еще неизвестны. Важное значение диметилирования H3K9 в сайленсинге гетерохроматиновых генов демонстрируется сильным зависящим от дозы влиянием SU(VAR)3-9 на PEV (обсуждается выше), атакже тем фактом, что супрессия сайленсинга генов мутациями Su(var)3-9 коррелирует с активностью их HKMT Ферментативно гиперактивная мутация Su(var)3-9 pth является сильным энхансером PEV и вызывает повышенное содержание H3K9me2 и H3K9me3 в хромоцентре, а также порождает заметные сигналы H3K9me2 во многих эухроматиновых сайтах (эктопический гетерохроматин). S-Аденозилметионин функционирует как донор метальной группы для всех этих реакций метилирования; следовательно, мутации в гене, кодирующем S-аденозилметионинсинтазу, Su(z)5, являются доминантными супрессорами PEV (Larsson et al., 1996).
Читать дальшеИнтервал:
Закладка:



