Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
1. Гены, оказывающиеся в ненормальном соседстве с гетерохроматином, обнаруживают мозаичный фенотип
Большие сегменты эукариотического генома, в основном повторяющиеся последовательности, упакованы в постоянно неактивной форме как конститутивный гетерохроматин. Эта фракция хроматина была исходно идентифицирована как та часть генома, которая остается конденсированной и интенсивно окрашивающейся (гетеропикнотической), по мере того, как клетка переходит от метафазы к интерфазе; такой материал обычно ассоциирован с теломерами и перицентромерными районами хромосом. Гетерохроматиновые районы обнаруживают тенденцию к поздней репликации и демонстрируют лишь незначительную мейотическую рекомбинацию или даже вовсе никакой. Эти районы бедны генами, но не лишены их вовсе, и те гены, которые там есть, для своей оптимальной экспрессии часто зависят от этой среды. Около одной трети генома Drosophila считается гетерохроматиновой, включая целую Y-хромосому, большую часть маленькой четвертой хромосомы, перицентромерные 40 % Х-хромосомы и перицентромерные 20 % больших аутосом. На протяжении нескольких последних десятилетий мы узнали очень многое о биохимии гетерохроматина, и многие из этих сведений получены в наших исследованиях, проведенных на Drosophila (Richards and Elgin, 2002; Schotta et al., 2003).
Одной из первых мутаций, идентифицированных у D. melanogaster, была мутация white — мутация, результатом которой оказывается белая окраска глаз мухи, а не характерная для них красная пигментация. Используя Х-лучи в качестве мутагена, Меллер (Muller, 1930) наблюдал необычный фенотип, при котором глаз был мозаичным, с участками красных и участками белых фасеток (рис. 5.1). Этот фенотип заставлял думать, что сам ген white не поврежден — в конце концов, некоторые фасетки оставались красными, и даже можно было получить мух-ревертантов с полностью красными глазами, вновь используя Х-лучи как мутаген. Однако ген white был несомненно сайленсирован в некоторых клетках, в которых он экспрессируется в норме Последующие исследования политенных хромосом (представлены ниже, см. рис. 5.4) показали, что такие фенотипы были следствием инверсии или перестройки, когда одна точка разрыва находилась в пределах перицентромерного гетерохроматина, а другая — вблизи гена white (см. рис. 5.1). Поскольку причина такого мозаичного фенотипа — в изменении положения гена в пределах хромосомы, это явление называется эффектом положения мозаичного типа (PEV) У Drosophila было показано, что по существу каждый ген, который был изучен в соответствующей перестройке, проявляет мозаипизм, и перестройки, включающие перицентромерный гетерохроматин любой хромосомы, могут приводить к PEV. PEV наблюдали у ряда организмов, в том числе у дрожжей, мух и млекопитающих, но как инструмент для изучения формирования гетерохроматина он был использован в основном у Drosophila.
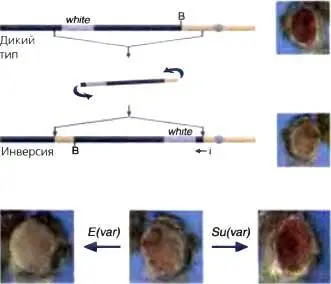
Рис. 5.1.Схематическое изображение мозаичности white в инверсии In(1)wm4 в Х-хромосоме
Локус white , в норме расположенный в дистальном эухроматине ( синий цвет ), теперь помещен в пределах 25 т.о. от точки разрыва в перицентромерном гетерохроматине ( розовый цвет ) Х-хромосомы благодаря индуцированной Х-лучами инверсии. Распространение гетерохроматиновой упаковки в эухроматиновый район приводит к сайленсингу; утрата сайленсинга в некоторых клетках в ходе дифференцировки дает мозаичный фенотип. Используя мух, обнаруживающих PEV, можно отселектировать мутации во втором сайте, которые либо супрессируют этот фенотип (мутации Su(var) ; приводят к утрате сайленсинга), либо усиливают фенотип (мутации Е(var) ; вызывают усиление сайленсинга)
PEV показывает, что такие перестройки позволяют упаковке в гетерохроматиновую конфигурацию «распространяться» вдоль по длине хромосомы. Все выглядит таким образом, как если бы перестройка удалила существующий в норме барьер или буферную зону. Следствием этого является измененная упаковка и сайленсинг генов, в норме организованных в эухроматиновой форме. Визуальное обследование политенных хромосом личинок, несущих такую перестройку, показывает, что район, несущий репортерный ген, упакован теперь в плотный блок гетерохроматина, но только в тех клетках, в которых этот ген неактивен (Zhimulev et al., 1986). Картина, наблюдаемая вследствие перестройки white, может варьировать по числу пигментированных клеток, величине пигментированных пятен и уровню содержания пигмента в наблюдаемых двух разных клеточных типах (рис. 5.1). В системе, использующей в качестве репортера индуцибельный ген lac-Z, исследователи наблюдали, что сайленсинг происходит в эмбриогенезе (когда впервые цитологически наблюдается гетерохроматин) и эпигенетически наследуется как в соматической, так и в зародышевой линиях; мозаичный фенотип был определен в ходе дифференцировки по мозаичной релаксации сайленсинга у личинок третьего возраста (Lu et al., 1996). Однако не все гены, обнаруживающие мозаицизм, остаются «молчащими» до периода после дифферениировки, и для разных генов баланс факторов, приводящий к решению «включить/ выключить», несомненно различен (дополнительное детальное обсуждение см. Ashburner et al., 2005b).
При наличии мух, имеющих PEV-фенотип, довольно просто провести скрининг на доминантные мутации во втором сайте (индуцированные химическими мутагенами, вызывающими точечные мутации или мелкие инсерции/делеции), которые либо являются супрессорами PEV (обозначаются Supressor of variegation , Su [varj) и приводят к утрате сайленсинга, либо являются энхансерами PEV (обозначаются Enhancer of variegation, E[var]), приводящими к усилению сайленсинга (рис. 5.1). Изолировали и охарактеризовали около 30 модификаторов PEV, но на основе такого скрининга можно предсказать существование значительно большего числа кандидатов на эту роль. Там, где такой ген был клонирован, а его продукт охарактеризован, обычно обнаруживают хромосомный белок или модификатор хромосомного белка (см. ниже). Небольшая выборка этих локусов вызывает как гаплоаномальный, так и триплоаномальный фенотип; т.е. если одна копия гена приводит к супрессии PEV, три его копии приводят к усилению PEV. Идентификация таких локусов привела к предположению, что белковые продукты этих генов играют структурную роль в гетерохроматине и что распространение гетерохроматиновой упаковки может управляться дозой этих белков в стохастическом режиме (рис. 5.2) (Locke et al., 1988). Однако «распространение» является не простым линейным континуумом, а сложным процессом, который, вероятнее всего, зависит от организации ДНК в том районе, который сайленсирован (см. ниже).
Читать дальшеИнтервал:
Закладка:



