Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Johnson L.M., Fisher-Adams G., and Gmnstein M. 1992. Identification of a non-basic domain in the histone H4 N-terminus required for repression of the yeast silent mating loci. EMBO J. 11: 2201-2209.
Johnson L.M., Kayne P.S., Kahn E.S., and Gmnstein M. 1990. Genetic evidence for an interaction between SIR3 and histone H4 in the repression of the silent mating loci in S. cerevisiae. Proc. Natl. Acad. Sci. 87: 6286-6290.
Kaeberlein M., McVey M., and Guarente L. 1999. The SIR2/3/4 complex and SIR2 alone promote longevity in S. cerevisiae by two different mechanisms. Genes Dev. 13: 2570-2580.
Kaeberlein M., Kirkland K.T., Fields S., and Kennedy B.K. 2004. Sir2-independent life span extension by calone restriction in yeast. PLoS Biol. 2: 296-307.
Kayne P.S., Kim U.J., Han M., Mullen J.R., Yoshizaki F., and Gmnstein M. 1988. Extremely conserved histone H4 N terminus is dispensable for growth but essential for repressing the silent mating loci in yeast. Cell 55: 27-39.
Kimura A., Umehara T., and Horikoshi M. 2002. Chromosomal gradient of histone acetylation established by Sas2p and Sir2p functions as a shield against gene silencing. Nat. Genet. 3: 370-377.
Kirchmaier A.L. and Rine J. 2001. DNA replication-independent silencing in S. cerevisiae. Science 291: 646-650.
Kobayashi X, Horiuchi T., Tongaonkar R, Vu L., and Nomura M. 2004. Sir2 regulates recombination between different rDNA repeats, but not recombination within individual rRNA genes in yeast. Cell 117: 441-453.
Laroche T., Martin S.G., Gotta M., Gorham H.C., Pryde RE., Louis E.J., and Gasser S.M. 1998. Mutation of yeast Ku genes disrupts the subnuclear organization of telomeres. Curr. Biol. 8: 653-656.
Lau A., Blitzblau H., and Bell S.P. 2002. Cell-cycle control of the establishment of mating-type silencing in S. cerevisiae. Genes Dev. 16: 2935-2945.
Li Y.C., Cheng T.H., and Gartenberg M.R. 2001. Establishment of transcriptional silencing in the absence of DNA replication. Science 291: 650-653.
Liou G.G., Tanny J.C., Kruger R.G., Walz T., and Moazed D. 2005. Assembly of the SIR complex and its regulation by O-acetyl-ADP-ribose, a product of NAD-dependent histone deacetylation. Cell 121: 515-527.
Loo S. and Rine J. 1994. Silencers and domains of generalized repression. Science 264: 1768-1771.
Luo K., Vega-Palas M.A., and Grunstein M. 2002. Rap1-Sir4 binding independent of other Sir, yKu, or histone interactions initiates the assembly of telomeric heterochromatin in yeast. Genes Dev. 12: 1528-1539.
Maillet L., Boscheron C, Gotta M., Marcand S., Gilson E., and Gasser S.M. 1996. Evidence for silencing compartments within the yeast nucleus: A role for telomere proximity and Sir protein concentration in silencer-mediated repression. Genes Dev. 10: 1796-1811.
Marcand S., Gilson E., and Shore D. 1997. A protein-counting mechanism for telomere length regulation in yeast. Science 275: 986-990.
Marcand S., Buck S.W., Moretti P., Gilson E., and Shore D. 1996. Silencing of genes at nontelomeric sites in yeast is controlled by sequestration of silencing factors at telomeres by Rap1 protein. Genes Dev. 10: 1297-1309.
Martin S.G., Laroche T., Suka N, Grunstein M., and Gasser S.M. 1999. Relocalization of telomeric Ku and SIR proteins in response to DNA strand breaks in yeast. Cell 97: 621-633.
Meneghini M.D., Wu M., and Madhani H.D. 2003. Conserved histone variant H2A.Z protects euchromatin from the ectopic spread of silent heterochromatin. Cell 112: 725-736.
Miller A.M. and Nasmyth K.A. 1984. Role of DNA replication in the repression of silent mating type loci in yeast. Nature 312: 247-251.
Mishra K. and Shore D. 1999. Yeast Ku protein plays a direct role in telomeric silencing and counteracts inhibition by Rif proteins. Curr. Biol. 9: 1123-1126.
Moazed D., Kistler A., Axelrod A., Rine J., and Johnson A.D. 1997. Silent information regulator protein complexes in S. cerevisiae: A S1R2/SIR4 complex and evidence for a regulatory domain in SIR4 that inhibits its interaction with SIR3. Proc. Natl. Acad. Sci. 94: 2186-2191
Moretti P, Freeman K., Coodly L., and Shore D. 1994. Evidence that a complex of SIR proteins interacts with the silencer and telomere-binding protein RAPl. Genes Dev. 8: 2257-2269.
Palladino E, Laroche X, Gilson E., Axelrod A., Pillus L., and Gasser S.M. 1993. SIR3 and SIR4 proteins are required forthe positioning and integnty of yeast telomeres. Cell 75: 542-555.
Pillus L. and Rine J. 1989. Epigenetic inheritance of transcriptional states in S. cerevisiae. Cell 59: 637-647.
Pryde F.E. and Louis E.J. 1999. Limitations of silencing at native yeast telomeres. EMBO J. 18: 2538-2550.
Renauld H., Aparicio O.M., Zierath P.D., Billington B.L., Chhablani S.K., and Gottschling D.E. 1993. Silent domains are assembled continuously from the telomere and are defined by promoter distance and strength, and by SIR3 dosage. Genes Dev. 7: 1133-1145.
Rusche L.N., Kirchmaier A. L., and Rine J. 2002. Ordered nucleation and spreading of silenced chromatin in Saccharomyces cerevisiae. Mol. Biol. Cell 7: 2207-2222.
-------- , 2003. The establishment, inheritance, and function of silenced chromatin in Saccharomyces cerevisiae. Annu. Rev. Biochem. 72: 481-516.
Sekinger E.A. and Gross D.S. 1999. SIR repression of a yeast heat shock gene: UAS and TATA footprints persist within heterochromatin. EMBO J. 18: 7041-7055.
Sinclair D.A. and Guarente L. 1997. Extrachromosomal rDNA circles—A cause of aging in yeast. Cell 91: 1033-1042.
Stavenhagen J.B. and Zakian V.A. 1994. Internal tracts of telomeric DNA act as silencers in Saccharomyces cerevisiae. Genes Dev. 8: 1411-1422.
Strahl-Bolsinger S., Hecht A., Luo K., and Grunstein M. 1997. SIR2 and SIR4 interactions differ in core and extended telomeric heterochromatin in yeast. Genes Dev. 11: 83-93.
Suka N., Luo K., and Grunstein M. 2002. Sir2p and Sas2p opposingly regulate acetylation of yeast histone H4 lysine 16 and spreading of heterochromatin. Nat. Genet. 3: 378-383.
Suka N., Suka Y., Carmen A.A., Wu J., and Grunstein M. 2001. Highly specific antibodies determine histone acetylation site usage in yeast heterochromatin and euchromatin. Mol. Cell 8: 473-479.
Taddei A., Hediger E, Neumann F.R., Bauer C, and Gasser S.M. 2004. Separation of silencing from perinuclear anchoring functions in yeast Ku80 Sir4 and Esc1 proteins. EMBO J. 23: 1301-1312.
Tanner K.G., Landry J., Stemglanz R., and Denu J.M. 2000. Silent information regulator 2 family of NAD-dependent histone/ protein deacetylases generates a unique product, 1-O-acetyl-ADP-ribose. Proc. Natl Acad. Set. 97: 14178-14182.
Thompson J.S., Johnson L.M., and Gmnstein M. 1994. Specific repression of the yeast silent mating locus HMR by an adjacent telomere. Mol. Cell. Biol. 14: 446-455.
van Leeuwen R, Gafken PR., and Gottschling D.E. 2002. Dotlp modulates silencing in yeast by methylation of the nucleosome core. Cell 109: 745-756.
Weiss K. and Simpson R.T. 1998. High-resolution stmctural analysis of chromatin at specific loci: S. cerevisiae silent mating type locus HMLa. Mol Cell. Biol. 18: 5392-5403.
Глава 5. Эффект положения мозаичного типа, формирование гетерохроматина и сайленсинг генов у Drosophila
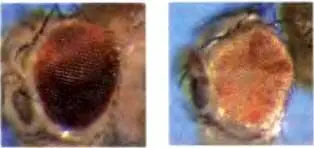
Sarah C.R. Elgin 1 и Gunter Reuter 2
1Department of Bilogy, Washington University, St. Louis, Missouri 63130
2Institute of Genetics, Biologicum, Martin Luther University Halle, D-06120 Halle, Germany
Общее резюме
Гены, оказавшиеся в ненормальном соседстве с гетерохроматином в результате либо перестройки, либо транспозиции [transposition], обнаруживают мозаичный фенотип, показывая тем самым, что данный ген оказался сайленсированным в некоторых клетках, в которых в норме он активен (эффект положения мозаичного типа — PEV, position-effect variegation). Сайленсинг, происходящий при PEV, можно приписать упаковке репортерного гена в гетерохроматиновой форме; это показывает, что формирование гетерохроматина, будучи однажды инициировано, может распространяться и охватывать близлежащие гены. На Drosophila melanogaster возможен генетический, цитологический и биохимический анализ, и в этой главе мы показываем, как эти разные подходы в совокупности помогли идентифицировать многих потенциальных участников этой системы, позволив в результате охарактеризовать несколько белков, играющих ключевую роль в установлении и поддержании гетерохроматина. Формирование гетерохроматина решающим образом зависит от метилирования гистона H3 по лизину 9, с сопутствующей ассоциацией Белка Гетерохроматина 1 (НР1 — Heterochromatin Protein 1) и других взаимодействующих белков, в том числе H3K9-метилтрансфераз; множественные взаимодействия этих белков необходимы для поддержания и распространения гетерохроматина. «Нацеливание» на гетерохроматин, в том числе накопление H3K9ше, происходит, по-видимому, с участием машинерии РНК-интерференции (RNAi), хотя играют роль и специфические ДНК-белковые взаимодействия. Хотя гетерохроматиновые районы (перицентромерные районы, теломеры и маленькая четвертая хромосома) обладают общей биохимией, каждый из них отличается от других, а перицентромерные районы мозаичны. Гетерохроматин у Drosophila беден генами, но не лишен их вовсе, и гены, находящиеся в гетерохроматине, зависят в своей экспрессии от этой среды. Окончательная модель формирования и поддержания гетерохроматина (включая «нацеливание» и распространение) должна будет принять в расчет различные реакции разных генов на это хроматиновое окружение.
Читать дальшеИнтервал:
Закладка:



