Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
10. Нарушение непрерывности репрессии естественных субтеломерных элементов теломерными петлями
Выше мы нарисовали упрощенную картину образования непрерывных участков «молчащего» хроматина, начинающихся с теломерных сайтов связывания белка Rap1. В действительности в природных теломерах все происходит более сложным образом в значительной степени из-за наличия пограничных элементов в области субтеломерных повторяющихся последовательностей. В большинстве экспериментов по включению в эту область репортерных конструкций для изучения теломерной репрессии субтеломерные повторяющиеся элементы X и Y’ удаляют. В результате репортерный ген оказывается непосредственно по соседству с TG-повторами. С другой стороны, все природные теломеры содержат субтеломерный повторяющийся элемент X, расположенный между TG-повторами и самым близким к теломере геном. Около 50—70% природных теломер содержат также не менее одной копии более длинного субтеломерного элемента У’ (рис. 4.8). И X, и У’ элементы содержат сайты связывания регуляторов транскрипции Tbf1 и Reb1, ограничивающие распространение «молчащего» хроматина (Fourel et al., 1999). Однако, X элементы содержат также каноническую последовательность ORC и сайты связывания Abf1, действующие противоположным образом. Они реинициируют или усиливают репрессию репортерных генов, находящихся с внутренней от них стороны. В результате сайленсинг в природных теломерах приобретает прерывистый характер, что отличает его от модели непрерывного распространения, изображенной на рис. 4.6. Для объяснения такого несовпадения Прайд и Луис (Pryde and Louis, 1999) также предложили модель петлеобразования в области теломеры, в которой область не репрессированного хроматина оказывается между двумя репрессированными доменами. Тем самым прерывистый характер «молчащих» доменов объяснялся без отказа от представлений о нуклеации и распространения гетерохроматина от TG-повторов.
11. Взаимодействие теломер in trans и перинуклеарное прикрепление гетерохроматина
Одной из наиболее консервативных черт гетерохроматина является его локализация в дискретных ядерных субкомпартментах. Это верно и для почкующихся дрожжей, у которых теломеры во время интерфазы образуют кластеры и остаются тесно ассоциированными с ядерной периферией. Такие кластеры были впервые обнаружены методом иммунно-окрашивания как фокусы локализации белков Rap1 и SIR на фоне их диффузного распределения по всему ядру (рис. 4.9). Подавление сайленсинга мутациями по остатку H4K16 или нарушениями в активности белков Rap1 и yKu сопровождается диспергированием белков SIR из этих кластеров (Hecht et al., 1995; Laroche et al., 1998). Позже было показано, что с ядерной оболочкой связаны не только теломеры, но и локусы HML и HMR Эта связь осуществляется несколькими дублирующими друг друга механизмами, зависящими либо от ассоциированного с тедомерой фактора yKu, либо от формирования «молчащего» хроматина как такового (Hediger et al., 2002). В «молчащем» хроматине функция прикрепления к ядерной оболочке приписывается субдомену белка Sir4, который связывается с белком ядерной оболочки Esc1 (enhances silent chromatin 1; Taddei et al., 2004). Взаимодействия Sir4-Esc1 прикрепляют SIR-репрессированные домены хроматина к участкам ядерной оболочки, отличным от ядерных пор. Даже в отсутствии прикрепления с помощью yKu-механизма ассоциация теломер с ядерной мембраной может поддерживаться посредством комплекса Sir4-Esc1 до тех пор, пока сохраняется сама репрессия. Более того, участки репрессированного хроматина, отделившиеся от теломер в результате рекомбинации, сохраняют SIR-зависимую ассоциацию с ядерной оболочкой (Gartenberg et al., 2004).
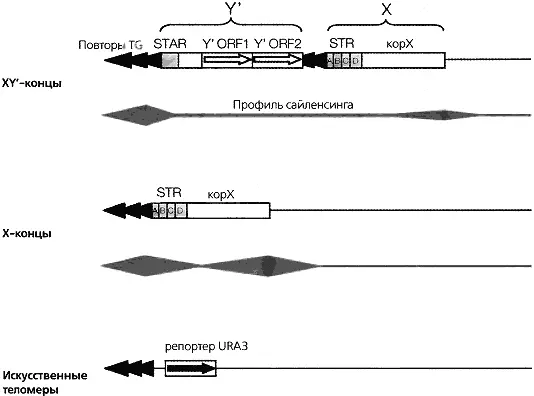
Рис. 4.8.Организация природных теломер и характер их сайленсинга
Показаны субтеломерные элементы и содержащиеся в них сайты связывания белков. Теломеры деляться на два основных класса: с Х-содержащими и Х+Y’-содержащими концами. Элементы STAR и STR подавляют распространение репрессии и оставляют область сниженной репрессии внутри Y’- и Х-элементов. Этого не наблюдается в искусственно укороченных теломерах, в которых присутствует градиент репрессии, распространяющийся на 3—4 т.п.н. от TG-повторов. Предполагается что природные теломеры образуют петли, аналогичные изображенным на рис. 4.6, в результате чего репрессированные области контактируют друг с другом, а не репрессированные остаются между областями контакта, (адаптировано из работы Pryde and Louis, 1999)
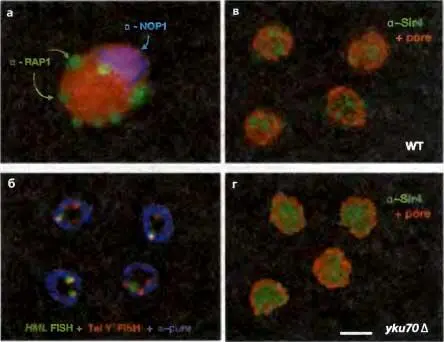
Рис. 4.9.SIR-белки и Rap1 образуют кластеры по периферии ядра
На панели а, Rap1 (зеленый) указывает на семь кластеров, представляющих все 64 теломеры данной диплоидной клетки Они расположены либо по периферии ядра, либо по соседству с ядрышком (голубое, помечено антителами к Nop1). ДНК помечена красным красителем. На панели б теломерная ДНК ( красная ) идентифицирована с помощью флюоресцентной гибридизации in situ (FISH), а HML помечен зеленым красителем. Они имеют одинаковую локализацию примерно в 70% случаев и находятся рядом с ядерной оболочкой (синяя) (Heun et al. 2001). Панель в показывает фокальное распределение Sir4 (зеленый) рядом с ядерной оболочкой (помечена МаЬ414, красная). Такой характер распределения утрачивается в штамме с делецией yKu 70, что совпадает с утратой теломерного сайленсинга (Laroche et al. 1998)
Исходно прикрепление теломер к ядерной оболочке, вероятно, происходит посредством yKu-зависимого пути, поскольку он работает даже в отсутствии сайленсинга. Благодаря этому прикреплению и взаимодействию между теломерами in trans происходит формирование ядерного субкомпартмента, который в свою очередь связывает SIR-белки (рис. 4.10). Этот компартмент имеет ключевое значение для образования градиента сайленсинга в области теломер. Даже фланкированные сайленсерами конструкты локуса НМ более эффективно репрессируются, когда интегрируются вблизи теломер (Thompson et al., 1994; Maillet et al., 1996) или когда они искусственно прикрепляются к ядерной оболочке трансмембранным фактором (Andrulis et al., 1998). Существенно, что способность усиливать репрессию за счет близости к теломерам утрачивается, когда Sir3 и Sir4 перестают захватываться кластерами или суперэкспрессируются (Maillet et al., 1996; Marcand et al., 1996). Это показывает, что градиент концентрации SIR-белков является той самой характеристикой теломерных кластеров, которая важна для усиления репрессии. Наконец, предполагается, что захват репрессоров общего назначения, присутствующих в лимитирующих концентрациях, помогает клетке обеспечить эпигенетическое наследование молчащего состояния, как показано на рис. 4.10. Вкратце, предложенная модель предполагает, что сборке вновь реплицированной ДНК в гетерохроматиновую структуру способствует ее локализация в субкомпартменте, обогащенном факторами сайленсинга.
Читать дальшеИнтервал:
Закладка:



