Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Следует заметить, что Sir1 участвует скорее в формировании гетерохроматиновой репрессии, чем ее поддержании. После установки репрессированного состояния участия Sir1 уже не требуется (Pillus and Rine, 1989). Его важная роль в установке репрессии была показана путем пришивки к нему ДНК-связывающего домена Gal4. При замещении сайленсера HMR- Е сайтами связывания Gal4 такой химерный GBD-Sir1 белок эффективно инициирует репрессию в отсутствии самого сайленсера и узнающего его фактора (Chien et al., 1993). Однако такая репрессия все равно требует участия остальных белков Sir и сохранности гистоновых хвостов. Следовательно одной из основных функций сайленсер-связывающих факторов является рекрутирование белка Sir1, который, в свою очередь, инициирует репрессию, рекрутируя другие Sir-белки во взаимодействия с близлежащими нуклеосомами. Это подтверждается и данными о том, что, в отличие от других Sir-белков, Sir1 не распространяется с комплексом SIR за пределы сайленсеров (рис. 4.5) (Rusche et al., 2002).
5.2. Теломерный гетерохроматин
В области теломер РНК-содержащий фермент теломераза поддерживает простую, но не регулярную TG-богатую повторяющуюся последовательность длиной в 300-350 н.п. Она содержит 16—20 сайтов связывания белка Rap1. Этот набор Rap1-связывающих сайтов образует ненуклеосомный «колпачек» на конце хромосомы и играет критическую роль в поддержании длины теломеры (Marcand et al., 1997). По длине теломерных повторов Rap1 связывается центральным ДНК-связывающим доменом со своими сайтами узнавания и С-концевым доменом с Sir4. Последнее происходит даже в отсутствие других SIR-белков и N-концов гистона Н4. Поскольку дефекты Sir4 нарушают связывание других белков с теломерным хроматином (Luo et al.. 2002), он, по-видимому, служит главным звеном между этапами нуклеации и следующим за ней формированием репрессированной структуры хроматина (рис. 4.6).
Связывающийся с концевыми участками ДНК комплекс yKu70/yKu80 помогает белку Rap1 рекрутировать Sir4 на концы хромосом. И действительно, утрата yKu резко уменьшает степень теломерной репрессии, а химерный белок GBD-yKu эффективно инициирует репрессию репортерных генов с поврежденными сайленсерными элементами. Необходимость yKu можно преодолеть элиминацией Rap1-связывающего фактора Rifl, конкурирующего с С-концевым доменом Rap1 за связывание с Sir4 (рис. 6) (Mishra and Shore, 1999). О кооперативных эффектах yKu и Rap1 в нуклеации гетерохроматина свидетельствует наблюдение, что 600-п.н. повторяющейся последовательности ДНК теломеры, содержащей более 30 сайтов связывания Rap1, недостаточно для нуклеации репрессии во внутренних локусах хромосом, хотя инсерпии 900-п.н. последователности, содержащей около 45 сайтов связывания Rap1, может быть достаточно (Stavenhagen and Zakian, 1994). Следует заметить, что в промоторах по всему геному дрожжей Rap1 служит транскрипционным фактором общего назначения. участвующим в активации многих генов, например, кодирующих белки рибосом. Почему в этих промоторах Rap1 рекрутирует факторы активации, а не нуклеации гетерохроматина, пока неизвестно.
6. Деацетилирование гистонов белком Sir2 обеспечивает сайты связывания для распространения SIR-комплексов
Молекулярные взаимодействия белков SIR хорошо изучены. Белок Sir4 играет роль ключевого «скелетного» фактора для их сборки. Sir4 и Sir2 активно взаимодействуют in vitro. Sir4 также независимо взаимодействует с Sir3, в то время как Sir3 и Sir2 взаимодействуют слабо (Moazed et al., 1997; Strahl-Bolsinger et al., 1997; Hoppe et al., 2002). Sir3 и Sir4 образуют также и гомодимеры (Moretti et al., 1994). При коэкспрессии в клетках насекомых Sir2, Sir3 и Sir4 образуют стабильный 360-kD комплекс, содержащий эти SIR-белки в стехиометрическом соотношении 1: 1: 1 (Cubizolles et al.. 2006). С моделью функционального гетеротримерного комплекса SIR2-3-4 согласуются и результаты исследования распределения этих трех белков в гетерохроматиновых доменах с помощью метода иммунопреципитации хроматина. Оказалось, что их содержание в разных участках гетерохроматиновых доменов одинаково (Hecht et al., 1996; Strahl-Bolsinger et al., 1997). Тем не менее, вполне очевидно, что белок Sir3 играет особую роль в распределении гетерохроматина. При суперэкспрессии Sir3 наблюдается расширение «молчащего» домена, совпадающее с распространением самого Sir3 за свою обычную границу в положении ~3 т.п.н. до -15 т.п.н. (Renauld et al., 1993; Hecht etal., 1996). Несбалансированная экспрессия одиночных Sir2 и Sir4 или даже их субдоменов имеет прямо противоположный эффект в виде нарушения ТРЕ, хотя скоординированная эктопическая экспрессия Sir3 и Sir4 противодействует дисбалансу и восстанавливает сайленсинг (Maillet et al., 1996). Все это иллюстрирует важное значение дозы белков комплекса SIR для его репрессорной функции, аналогично ситуации с комплексами Polycomb у дрозофилы. Уникальная способность Sir3 распространяться вдоль доменов хроматина при суперэкспрессии коррелирует с наблюдением, что in vitro он образует стабильные мультимеры (Liou et al., 2005).
Основа, по которой распространяется SIR-комплекс, представляет собой нуклеосомы с деацетилированными по N-концевым участкам гистонами H3 и Н4 (Braun-stein et al., 1996; Suka et al., 2001). Механизм распространения можно объяснить способом взаимодействия SIR-белков с гистонами (рис. 4.7). Белки Sir3 и Sir4 связываются с деацетилированными N-концами гистонов H3 и Н4 in vitro и in vivo (Hecht et al., 1995, 1996), причем участия хвостов H2A и Н2В для этого взаимодействия не требуется. Наиболее важная в этом отношении область гистонов — 16—29-й аминокислотные остатки гистона Н4, среди которых 16-й остаток лизина, в частности, должен быть деацетилирован (положительно заряжен) для связывания Sir3 (Johnson et al., 1990,1992). В отличие от мутаций по другим сайтам ацетилирования, даже консервативные мутации по остатку H4K16 полностью нарушают теломерный сайленсинг. Хвосты гистонов H3/Н4, и особенно область 16—24-го остатков Н4, способствуют компактизации нуклеосомных цепей in vitro , а деацетилирование H4K16 в этом случае, вероятно, регулирует наднуклеосомные уровни сворачивания нуклеосомной цепи. Как же регулируется само деацетилирование H4K16 in vivo ?
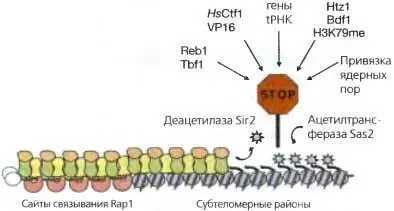
Рис. 4. 7. Пограничные функции гетерохроматина у почкующихся дрожжей
Распространение гетерохроматина посредством деацетилирования белком Sir2 гистона Н4 по остатку К16 ограничивается конкурирующей активностью гистоновой ацетил-трансферазы Sas2, ацетилирующей H4K16 в прилегающем эухроматине, тем самым предотвращая связывание Sir3. Метилирование H3K79 в прилегающем эухроматине также влияет на распространение гетерохроматина. Пограничные функции могут иметь и такие факторы как Reb1, Tbf1 и белки млекопитающих и вирусов Ctf 1 и VP 16, прикрепление к ядерным порам и присутствие генов тРНК. Вполне возможно, что некоторые из них действуют, рекрутируя гистоновые ацетилтрансферазы
Читать дальшеИнтервал:
Закладка:



