Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Рис. 4.3.Переключение типов спаривания у дрожжей
(а)Гомоталлические штаммы дрожжей способны переключать тип спаривания с каждым клеточным циклом. Переключение происходит до репликации ДНК, поэтому и материнская, и дочерняя клетки приобретают новый тип спаривания.
(б)Показано положение «молчащего» и экспрессирующегося локусов типа спаривания на III хромосоме. Активный локус МАТ может переключаться посредством конверсии генов примерно один раз на каждый клеточный цикл, благодаря двунитчатым разрывам ДНК эндонуклеазой НО. Указанные проценты показывают частоту, с которой события конверсии переключают локусы МАТ между противоположными типами. Направленность переключения обеспечивается энхансером рекомбинации (RE) на левом плече III хромосомы.
(в)Репрессия «молчащих» локусов типов спаривания HMR и HML происходит под действием двух сайленсеров, фланкирующих эти гены. Они называются Е (от essential) и I (от important) (Brand et al., 1997) и содержат участки связывания белков Rap1 (R), Abf1 (А) и ORC (О). Комбинируя такие сайты связывания в разных комбинациях, можно создавать искусственные сайленсеры, хотя их эффективность ниже, чем у природных. НМL α и HMRa расположены на расстоянии 12 и 23 т.п.н. от теломеры III хромосомы, соответственно. Домены теломерного гетерохроматина III хромосомы репрессируются независимо от локусов НМ процессом, инициируемым множественными сайтами связывания Rap1 (R)
Гетерохроматин присутствует также непосредственно по соседству с повторами теломерной ДНК дрожжей (C 1-3A/TG 1-3). Как упоминалось выше, при интеграции репортерных генов, таких как Ura3 или Ade2, рядом с теломерными повторами происходит их репрессия по мозаичному и эпигенетическому типу (Gottschling et al. 1990) Этот эффект (ТРЕ), как и сайленсинг локусов НМ, зависит от Rap1, Sir2, Sir3, Sir4 и N-концевых участков молекул гистонов (Kayne et al., 1988; Aparicio et al., 1991). Генетические исследования свидетельствуют о том, что, за исключением Sir1 , близкие механизмы обеспечивают сайленсинг в области локусов НМ и в примыкающих к теломерам участках. Более того, учитывая способность репортерных генов в субтеломерных участках достаточно часто переключаться между «молчащим» и экспрессирующимся состояниями, данный тип репрессии, по-видимому, весьма близок явлению мозаичности, обусловленной эффектом положения (PEV) у дрозофилы.
У дрожжей четыре белка Sir, осуществляющих репрессию, не имеют существенной взаимной гомологии. Белки Sir1, Sir3 и Sir4 консервативны только у самих S. cerevisiae и близких им видах почкующихся дрожжей. Sir2, напротив, является родоначальником большого семейства НАД-зависимых деацетилаз гистонов, консервативного от бактерий до человека (рис. 4.4). Роль 8п2-подобных деацетилаз гистонов в репрессии транскрипции обнаружена даже у организмов, не имеющих других Sir белков, таких как делящиеся дрожжи и двукрылые. У дрожжей Schizosaccharomyces pombe активность Sir2 необходима для сайленсинга транскрипции вблизи теломер, а у дрозофилы она влияет на стабильность эффекта PEV (см. обзор Chopra and Mishra, 2005). Сопряженный с деацегилированием гидролиз НАД белком Sir2 приводит к образованию О-ацетил-АДФ-рибозы, интермедиата, возможно имеющего свою собственную функцию (Tanner et al. 2000; также см. раздел 13). Важно отметить, что помимо гистонов ферменты семейства Sir2 модифицируют множество других субстратов, причем имеется обширная ветвь этого семейства, состоящая из цитоплазматических ферментов (рис. 4.4). Разнообразие функций Sir2 можно проиллюстрировать тем, что у млекопитающих ферменты этого семейства деацетилируют факторы транскрипции FOXO и р53 в ответ на стресс и повреждение ДНК, изменяя тем самым их взаимодействие. У почкующихся дрожжей Sir2 имеет важную роль помимо участия в сайленсинге, а именно подавляет нереципрокную рекомбинацию высокоповторяющихся генов рДНК локуса в области ядрышка (Gottlieb and Esposito 1989).
4. Гетерохроматин отличается репрессивной структурой, которая распространяется на весь «молчащий» домен
Репрессия активности генов в эухроматине может происходить вследствие связывания репрессорного белка или комплекса, узнающего определенную последовательность в промоторе гена и препятствующего движению или посадке аппарата транскрипции. Репрессия в области гетерохроматина достигается другим способом, который не является промотор-специфичным. Она начинается с определенных участков, от которых распространяется непрерывно по всему домену, репрессируя все промоторы, находящиеся в соответствующей области (рис. 4.5) (Renauld et al., 1993). Наиболее четко это было показано с помощью метода иммуно-преципитации хроматина: оказалось, что белки Sir2, Sir3 и Sir4 физически связаны с хроматином по всему «молчащему» субтеломерному домену (Hecht et al., 1996; Strahl-Bolsinger et al., 1997). Доказательства, что такая связь приводит к формированию менее доступной репрессивной структуры были получены в других экспериментах. Например, ДНК «молчащих» доменов хроматина плохо метилировалась экспрессируемой в дрожжевых клетках бактериальной метилазой dam , тогда как последовательности ДНК вне этих доменов метилировались вполне эффективно. По-видимому, гетерохроматин затрудняет доступ к ДНК таким молекулам как dfom-метилтрансфераза (Gottschling. 1992). Аналогично, 3-т.п.н. локус HMR избирательно устойчив к действию определенных рестрикционных эндонуклеаз в изолированных ядрах (Loo and Rine, 1994), а нуклеосомы между двух сайленсер-элементов в «молчащих» локусах НМ расположены близко друг к другу, образуя нуклеазо-резистентный домен, отсутствующий в активных локусах (Weiss and Simpson, 1998). Итак, гетерохроматин у дрожжей имеет особую структурную организацию.
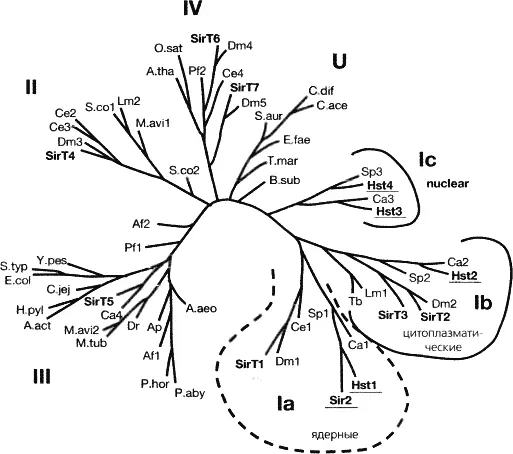
Рис. 4.4.Семейство деацетилаз Sir2
Sir2 является прототипом большого семейства НАД-зависимых деацетилаз. Семейство белков Sir2 необычно консервативно и найдено у организмов от бактерий до человека. Его эволюционное древо содержит как ядерные, так и цитоплазматические ветви. Это филогенетическое древо без корня было построено с помощью программ CLUSTAL W® и TREEVIEW® для сравнения последовательности коровых доменов гомологов, идентифицированных в библиотеках кДНК и уникальных генов. Шесть подклассов и несвязанная группа описаны в работе Фрая (Frye 2000). Гомологи млекопитающих обозначены как SirT1-7 жирным шрифтом, а белки почкующихся дрожжей подчеркнуты Остальные обозначены названием вида (модифицировано с разрешения из Frye 2000 [©Elsevier])
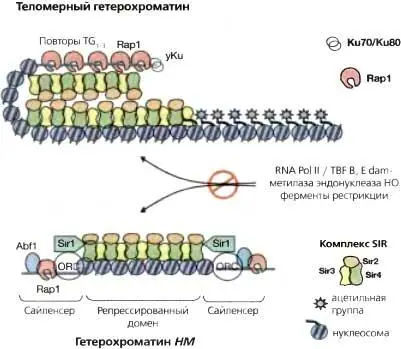
Рис. 4.5.Модель гетерохроматина дрожжей в теломерах и локусах НМ
Читать дальшеИнтервал:
Закладка:



