Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Для распространения SIR-комплексов механизмы сайленсинга и теломер, и НМ- локусов используют Rap1, Sir2, Sir3 и Sir4, но они различаются тем, что теломерный зависит также от yKu, в то время как НМ- сайленсеры используют факторы ORC, Abf1 и Sir1. Предполагается, что теломерный гетерохроматин образует обратную петлю сам на себя. В результате получается структура, защищающая теломеру от деградации. Ее конденсация и сворачивание приводят к сайленсингу генов. В случае HM -гетерохроматина репрессированный домен между сайленсерными элементами состоит из тесно расположенных нуклеосом, образующих конденсированную структуру И теломерные, и НМ «молчащие» области недоступны аппарату транскрипции и деградирующим ферментам
Не столь ясен вопрос о степени гиперконденсации гетерохроматина у дрожжей и многоклеточных организмов, и обусловленного ею затруднения доступа факторов транскрипции к ДНК. Репрессивный комплекс, образующийся при взаимодействии белков Sir с гистонами, оказался неожиданно динамичным: эти белки могут включаться в хроматин «молчащего» локуса НМ даже при остановке клеток в фазе клеточного цикла, в которой образования гетерохроматина обычно не происходит (Cheng and Gartenberg, 2000). Возможно, именно этим обстоятельством объясняется способность Sir-содержащего хроматина связываться с определенными факторами транскрипции даже в репрессированном состоянии (Sekinger and Gross, 1999). Тем не менее, хотя эти исследования и свидетельствуют о том, что гетерохроматин не является механическим препятствием для связывания с ДНК всех негистоновых белков, в нем не обнаруживается транскрипционной активности и связанных РНК-полимераз. Судя по результатам экспериментов Чена и Видома (Chen and Widom, 2005), специфически подавляемым гетерохроматином процессом является связывание с промотором комплексов РНК-полимеразы II и транскрипционных факторов TFIIB и TFIIE. Итак, хотя «молчащий» хроматин дрожжей является динамичным образованием, допускающим обмен белков SIR и, возможно, некоторых факторов транскрипции, он избирательно препятствует связыванию основного аппарата транскрипции и тем самым блокирует синтез мРНК (более детально см. главу 10).
5. Отдельные этапы сборки гетерохроматина
Формирование гетерохроматина у всех видов включает несколько молекулярных этапов, некоторые из которых были найдены и у почкующихся дрожжей. Один из наиболее изученных из них — сайт-специфическая нуклеация гетерохроматина, требующая участия сайт-специфических ДНК-связывающих факторов. Затем гетерохроматин распространяется из сайтов инициации. Распространение ограничивается специальными пограничными механизмами, которые у дрожжей хорошо изучены. Наконец, дрожжи успешно использовали для демонстрации роли субъядерных компартментов в опосредованной гетерохроматином репрессии. Формирование гетерохроматина в области теломер несколько отличается от его формирования в локусах НМ , но в обоих случаях действует общий принцип, что специфические ДНК-связывающие факторы инициируют распространение репрессоров общего назначения. В деталях эти механизмы описаны ниже (рис. 4.6).
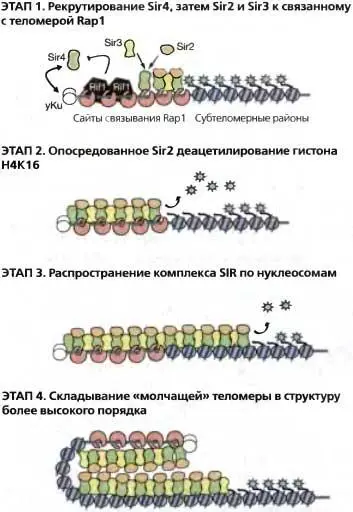
Рис. 4.6.Этапы сборки теломерного гетерохроматина
ЭТАП 1. Рекрутирование Sir4, затем Sir2 и Sir3 связанным с теломерами Rap1. Нателомерах Rap1 и yKuрекрутируют Sir4даже в отсутствии Sir2 и Sir3. Только Sir4 может быть рекрутирован в отсутствии остальных Sir-белков, причем его связыванию препятствуют Rifl и Rif2 (Mishra and Shore 1999).
ЭТАП 2. Опосредованное Sir2 деацетилирование гистона H4K16. Sir4-Sir2 и Sir4-Sir3 активно взаимодействуют, образуя Sir-комплексы по длине TG-повторов. НАД-зависимая гистон-деацетилазная активность Sir2 стимулируется образующимися комплексами и деанетилирует К16 остаток гистона Н4 в близлежащих нуклеосомах.
ЭТАП 3. Распространение SIR-комплекса по нуклеосомам. SIR-комплексы распространяются по нуклеосомам, возможно, с использованием О-ацетил-АДФ-рибозных интермедиатов, образующихся в результате НАД-гидролиза (Liou et al. 2005). Sir3 и Sir4 связываются с деацетилированными хвостами гистона Н4. Хотя деацетилированный N-концевой хвост гистона H3 также связывает белки Sir3 и Sir4, он не показан.
ЭТАП 4. Свертывание « молчащей» теломеры в структуру более высокого уровня. «Молчащий» хроматин «созревает» в конце М-фазы с образованием недоступной структуры. Этот процесс может включать более высокие уровни свертывания и связывание с ядерной оболочкой.
5.1. НМ гетерохроматин
«Молчащие» локусы типов спаривания HML и HMR ограничены по бокам короткими последовательностями ДНК — сайленсерами, обозначаемыми как Е (essential — существенный) и I (important — важный; рис. 4.36, в).
Сайленсеры содержат участки связывания, как минимум, двух полифункциональных ядерных факторов, а именно Rap1 и Abf1, а также ориджин-узнающего комплекса (origin recognition complex — ORC) (Brand et al., 1987). Хотя делепия HMR-E, содержащего все три участка узнавания, гораздо сильнее влияет на сайленсинг, чем делеция HMR-1 , содержащего лишь два из них, каждый из сайленсеров в локусах HMR и HML может служить специфическим сайтом нуклеации для связывания репрессирующего Sir-комплекса и последующего распространения белков Sir на лежащие между сайленсерами нуклеосомы. Показано, что между молекулами Rap1, ассоциированными с разными участками связывания in vitro, существуют непосредственные физические контакты. Это свидетельствует о том, что факторы, ассоциированные с сайленсерами Е и I, могут взаимодействовать друг с другом путем «выпетливания» лежащего между ними репрессированного домена. Такая модель легко объясняет наличие кооперативного эффекта этих сайленсеров на инициацию репрессии (Hofmann et al., 1989).
Избыточность функции сайленсерных элементов является характерной чертой гетерохроматиновой репрессии и наблюдается также внутри самих сайленсеров. Факторы транскрипции общего назначения Rap1 и Abf1, а также ориджин-узнающий комплекс ORC, обеспечивающий посадку пререпликативного комплекса на ориджины репликации, функционально дублируют друг друга. Как показали эксперименты с делециями, сайтов связывания любых двух из этих факторов достаточно для сайленсинга (Brand et al., 1987), хотя сколько-нибудь существенного структурного сходства между Rap1, Abf1 и ORC не обнаружено. Их функциональная избыточность объясняется рекрутируемыми (вовлекаемыми) ими белками. Например, Rap1 рекрутирует белок Sir4 в области сайленсеров НМ и теломер, Abf1 взаимодействует с Sir3, а ORC имеет высокое сродство к Sir1, специфическому для репрессии локусов НМ фактору SIR (см. обзор Rusche et al., 2003). Сам Sir1 непосредственно взаимодействует с N-концом Sir4, образуя мостик между ORC и комплексом SIR2-3-4. Таким образом, разные связывающиеся с сайленсерами факторы приводят к рекрутированию Sir4, а затем и комплекса SIR2-3-4, необходимого для репрессии во всех случаях. Видимая избыточность между Rap1, Abf1 и ORC (в сайленсерах), а также гетеродимера Ки (в области теломер, см. ниже), может быть связана с их способностью инициировать репрессию с помощью прямых контактов с разными компонентами комплекса SIR.
Читать дальшеИнтервал:
Закладка:



