Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Геном N. crassa (~40 млн о.) состоит из семи хромосом с примерно 10000 предсказанными генами, кодирующими белки (Galagan et al., 2003), и общей длиной генетической карты, равной, огрубленно, 1000 единицам (Perkins et al., 2001). Лишь около 10 % этого генома состоят из повторяющейся ДНК, и, помимо тандемного порядка из -70 копий сегмента рДНК длиной ~9 т.н., кодирующего три большие рРНК, большая часть повторяющейся ДНК состоит из инактивированных перемещающихся элементов. Тот факт, что большинство штаммов Neurospora не имеет активных транспозонов и имеет очень мало близких паралогов, почти наверняка отражает действие RIP, первой открытой у эукариот системы защиты генома, зависящей от гомологии (Selker, 1990). Мы знаем, что у Neurospora имеются по крайней мере три процесса сайленсинга генов, которые должны служить для консервации структуры генома: RIP, подавление (quelling) и MSUD (Borkovich et al., 2004). У всех этих процессов имеются эпигенетические аспекты и все они имеют прямые или косвенные связи с метилированием ДНК — базовым эпигенетическим механизмом, обнаруживаемым у Neurospora и многих других эукариот. Мы обсудим метилирование ДНК, а затем RIP, подавление (quelling) и MSUD.
2.1. Метилирование ДНК у Neurospora
Со времени своего открытия десятилетия тому назад метилирование ДНК у эукариот остается в высшей степени загадочным. Все еще вызывают споры такие базовые вопросы, как вопрос о том, что определяет, какие именно хромосомные районы метилируются, и какова функция метилирования ДНК. Neurospora проявила себя как превосходная система для изучения контроля и функции метилирования ДНК. У некоторых модельных эукариот, в том числе нематоды С. elegans и дрожжей S. cerevisiae и
S. pombe, сколько-нибудь заметное метилирование ДНК отсутствует, а сообщения о метилировании ДНК у еще одного модельного организма, D. melanogaster, остаются спорными. У некоторых организмов, таких как млекопитающие, метилирование ДНК имеет существенное значение для выживаемости, усложняя некоторые анализы. В ДНК Neurospora около 1,5 % цитозинов метилированы, но этим метилированием можно пренебречь, что облегчает генетические исследования. Хотя следует быть осторожным при экстраполяции с одной системы на другую, по крайней мере некоторые аспекты метилирования ДНК оказываются консервативными. Например, все известные ДНК-метилтрансферазы (DMTs), включая DMTs и прокариот, и эукариот, обнаруживают поразительную гомологию в своих каталитических доменах (Grace Goll and Bestor, 2005). Полученные за последнее десятилетие данные по Neurospora, Arabidopsis, мышам и другим системам выявили как важное сходство, так и интересные различия в контроле и функциях метилирования ДНК, демонстрируя ценность проведения исследований на многих модельных системах.
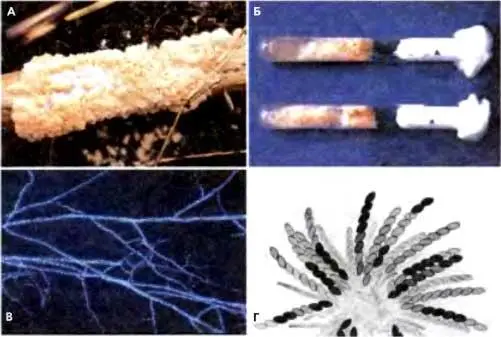
Рис. 6.10.Изображения N. crassa
(А) Вегетативный рост в природных условиях на сахарном тростнике (фото D. Jackobson; Stanford University)
(Б) Агаровые «косяки» с вегетативными культурами N. crassa в лаборатории (фото N.B.Raju, Stanford University)
(В) Гифы N. crassa, окрашенные DAPI, чтобы показать многочисленные ядра (фото М. Springer, Stanford University).
(Г) Розетка созревающих асков, демонстрирующая разные паттерны аскоспор. (Г перепечатано, с любезного разрешения, из: Raju, 1980 [©Elsevier])
Открытие метилирования ДНК у Neurospora первоначально вызвало интерес в связи с тем, что оно не было ограничено симметричными сайтами, такими как динуклеотиды CpG или тринуклеотиды CpNpG. Риггс (Riggs), а также Холлидей и Нью (Holliday и Pugh), предложили привлекательную модель для «наследования» или «поддержания» паттернов метилирования, базирующуюся на симметричной природе метилированных сайтов у животных. Хотя результаты разнообразных исследований in vitro и in vivo были в пользу модели «поддерживающей метилазы» (глава 18), можно представить себе и механизмы поддерживающего метилирования, которые не основываются на точном копировании в симметричных сайтах; такие механизмы могут действовать у ряда организмов (см., например: Selker et al., 2002). Вероятность того, что наблюдаемое метилирование в асимметричных сайтах представляло собой «метилирование de novo», казалась очень привлекательной, потому что механизмы, слепо воспроизводящие паттерны метилирования, могут усложнять определение нуклеотидных последовательностей, метилируемых изначально [in the first place]. Действительно, проведенные на Neurospora исследования с ДНК-опосредованной трансформацией и ингибиторами метилирования продемонстрировали воспроизводящееся метилирование de novo (см., например: Singer et al., 1995). Дополнительные исследования определили, отчасти, основные сигналы для метилирования de novo (см., например: Tamaru and Selker, 2003).
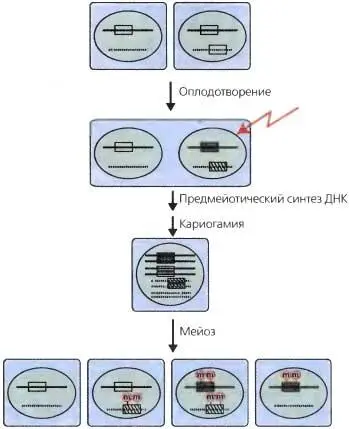
Рис. 6.11.Индуцированная повторами точечная мутация (RIP)
Для упрощения показаны лишь две хромосомы. Пустой прямоугольник представляет ген или любой хромосомный сегмент, который во время дупликации (например, в штамме, показанном наверху справа ) подвержен R1P (символизируется знаком молнии ) между оплодотворением и кариогамией. Результаты генетических экспериментов показывают, что дупликации могут повторно подвергаться залпам транзиций С в Т (символизируются заполненными прямоугольниками ) на протяжении этого периода времени, равного -10 митозам, как раз к конечному премейотическому синтезу ДНК [right up to the final premeiotic DNA synthesis] (Selker et al., 1987; Watters et al., 1999). Показаны четыре возможные комбинации хромосом в потомстве, и красное «т» изображает метилирование ДНК, которое часто (хотя и не всегда) связано с продуктами RIP
Первым метилированным пэтчем, охарактеризованным в деталях, был участок длиной 1,6 т.о., который состоит из дивергировавшей тандемной дупликации сегмента ДНК длиной 0,8 т.о., включающего ген 5S рРНК. Сравнение этого участка с соответствующим хромосомным районом таких штаммов, у которых этой дупликации нет, первоначально привело к идее, что повторяющиеся последовательности могут каким-то образом индуцировать метилирование ДНК, и в конце концов привело к открытию системы защиты генома, названной RIP. Расшифровка RIP показала, что повторяющиеся последовательности не могут прямо включать метилирование ДНК, по крайней мере у Neurospora ; вместо этого повторы включают RIP, который тесно связан с метилированием ДНК, как описано ниже. И район ?-?, и район ?63, второй метилированный район, открытый у Neurospora , являются продуктами RIP. Более того, последующие анализы метилирования ДНК по всему геному обнаружили, что большинство метилированных участков у Neurospora являются реликтами транспозонов, инактивированных RIP (Galagan et al., 2003; Selker et al., 2003). Действительно, единственное метилирование ДНК у Neurospora , которое , возможно, не является результатом RIP, — это метилрование в тандемных порядках рРНК (Perkins et al., 1986).
Читать дальшеИнтервал:
Закладка:



