Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
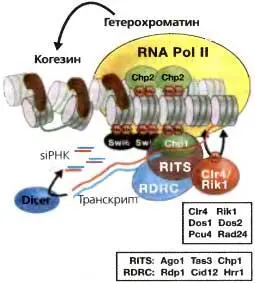
Puc. 6.6.Транскрипция центромерных повторов связывает RNAi, образование гетерохроматина и когезию
Транскрипция внешних повторов с помощью RNA pol II обеспечивает первичный субстрат для RNAi и зависимого от Dicer образования siRNA. Загрузка Ago1 в комплексе RITS (Ago1, Tas3, Chp1) молекулами siRNA делает возможным «нацеливание» гомологичного транскрипта. Действие RDRC (Rdpl, Cidl2 и Hrrl) делало бы возможной продукцию dsRNA, обеспечивая большее количество субстрата для Dcr1 для производства siRNA и, возможно, усиления сигнала. Взаимодействия между транскриптом, субъединицами RNA pol II и компонентами RNAi рекрутируют Clr4, который метилирует гистон H3 по лизину 9, делая возможным связывание хромодоменных белков, рекрутирование когезина и когезию сестринских центромер
Мутанты, вызывающие утрату сайленсинга во внешних повторах (т.е. дефектные по Clr4, компонентам RNAi или Swi6), повысили частоту утери хромосом в митозе и встречаемость отстающих хромосом на веретенах поздней анафазы (рис. 7а) (Allshire et al., 1995; Ekwall et al., 1996; Bernard et al., 2001; Nonaka et al., 2002; Hall et al., 2003; Volpe et al., 2003). Клетки без Swi6 дефектны по когезии в центромерах, но сохраняют когезию вдоль хромосомных плеч (Bernard et al., 2001; Nonaka et al., 2002). Формирование должным образом би-ориентированного веретена требует, чтобы сестринские кинетохоры прикреплялись к микротрубочковым фибрилам, отходящим от противоположных полюсов веретена. Силы, прилагаемые к би-ориентированным кинетохорам, требуют, чтобы сестринские кинетохоры прочно удерживались вместе (рис. 6.76). Swi6 необходим для рекрутирования когезина к хроматину внешних повторов и, тем самым, опосредует прочную физическую когезию между сестринскими центромерами (рис. 6.6). Таким образом, одна из функций «молчащего» хроматина в центромерах — быть посредником в когезии.
Когезин также прочно ассоциирован с теломерами и районом mat2-mat3 (Bernard et al., 2001; Nonaka et al., 2002). Кроме того, когезин также рекрутируется к «молчащему» хроматину, сформированному на гене ura + в ответ на расположенный рядом эктопический центромерный повтор (рис. 6.5), подчеркивая связь между «молчащим» хроматином и когезией (Partridge et al., 2002). Таким образом, рекрутирование когезина, по-видимому, является общим свойством ассоциированного со Swi6 «молчащего» хроматина. Каким образом Swi6-хроматин осуществляет рекрутирование когезина — неизвестно, но Swi6 взаимодействует с субъединицей Psc3 когезина (Nonaka et al., 2002). Кроме того, Dfp1, регуляторная субъединица консервативной киназы Hskl (Cdc7), взаимодействует с Swi6 и требуется для рекрутирования когезина к центромерам (Bailis et al, 2003). Эта функциональная связь между гетерохроматином и когезией хроматид, по-видимому, сохраняется и у других организмов, поскольку уменьшение содержания компонента RNAi, Dicer, по-видимому, влияет на целостность гетерохроматина и когезию сестринских центромер в клетках позвоночных (Fukagawa et al., 2004).
1.8. Эпигенетическое наследование функционального состояния центромеры
Интересное эпигенетическое явление было описано в отношении сборки функциональных центромер у дробянковых дрожжей на плазмидах, содержащих минимальные участки для центромерной функции. Хотя конструкты, сохраняющие лишь часть внешнего повтора и большую часть центрального домена, собирают функциональную центромеру неэффективно, но, как это ни удивительно, коль скоро это активное функциональное состояние центромеры установлено, оно может воспроизводиться в ряду многих митотических делений и даже в мейозе (Steiner and Clarke, 1994; Ngan and Clarke, 1997). Одно из объяснений этого сводится к тому, что внешние повторы обеспечивают среду, благоприятную для сборки кинетохора (Pidoux and Allshire, 2005), но, будучи собранным, CENP-A Cnp1-хроматин (и, следовательно, кинетохор) воспроизводится в этом положении с помощью матричного механизма, который может быть сопряжен с репликацией (Takahashi et al., 2005). Возможно, что гетерохроматин каким-то образом индуцирует откладку CENP-A Cnp1в центральном домене или способствует ей (рис. 6.8) и что один только блок гетерохроматина не обеспечивает эффективную сборку кинетохора. Альтернативное объяснение заключается в том, что один внешний повтор не достаточен для рекрутирования достаточных количеств когезина, и это приводит к дефектной центромерной когезии и повышенным частотам утери хромосом. Такие центромерные конструкты после нескольких клеточных делений могут стохастически накапливать достаточные количества когезина, результатом чего оказывается увеличенная митотическая стабильность. Однажды достигнутое, это стабилизированное состояние должно каким-то образом дуплицироваться на дочерних молекулах, чтобы сделать возможным его воспроизведение в последующих делениях (рис. 6.8).
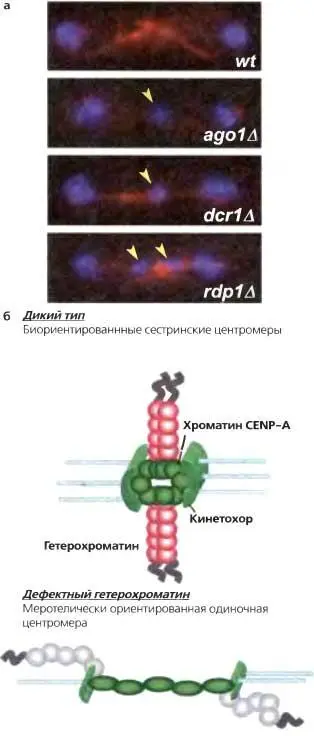
Рис. 6.7.Утрата гетерохроматина приводит к дефектному расхождению хромосом
(а) Клетки, лишенные RNAi или компонентов гетерохроматина, обнаруживают увеличенную частоту хромосомных потерь и отстающих хромосом на веретенах в поздней анафазе, (б) Отстающие хромосомы в клетках с дефектным гетерохроматином могут быть результатом дезорганизации кинетохоров, так что одна центромера может прикрепляться к микротрубочкам от противоположных полюсов. Такая меротелическая ориентация могла бы сохраняться при переходе в анафазу; разрыв прикрепления к одному или другому полюсу приводил бы к случайной сегерегации, результатом чего были бы случаи потери-приобретения хромосом
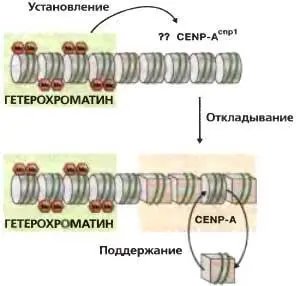
Рис. 6.8.Установление и поддержание хроматина CENP-A
ДНК центрального домена сама по себе не способна устанавливать функциональную центромеру; необходимы внешние повторы. Утрата гетерохроматина из установленных центромер не влияет на сборку CENP-ACnp1 или кинетохора на центральном домене. Это позволяет предполагать, что гетерохроматин каким-то образом указывает сайт CENP-ACnp1-хроматина и, тем самым, сборки кинетохора. Неизвестно, как CENP-ACnp1-хроматин откладывается в нуклеосомах или каким образом этот специализированный хроматин поддерживается в центральном домене
Как упоминалось выше, TSA также может устанавливать дефектное по центромере состояние, вызываемое утратой сайленсинга (Ekwall et al., 1997). С учетом связи между формированием «молчащего» хроматина и когезией в центромерах представляется вероятным, что индуцированное TSA гиперацетилирование блокирует эффективную реконструкцию «молчащего» хроматина посредством RNAi и что дефектная функция центромеры воспроизводится благодаря утрате сайленсинга и, таким образом, когезии сестринских центромер (дополнительные детали см. в главе 14).
Читать дальшеИнтервал:
Закладка:



