Роберт Сапольски - Биология добра и зла. Как наука объясняет наши поступки

- Название:Биология добра и зла. Как наука объясняет наши поступки
- Автор:
- Жанр:
- Издательство:Альпина нон-фикшн
- Год:2019
- Город:Москва
- ISBN:978-5-0013-9051-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Роберт Сапольски - Биология добра и зла. Как наука объясняет наши поступки краткое содержание
Биология добра и зла. Как наука объясняет наши поступки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Таким образом, у нас есть множество классов нейромедиаторов, и каждый связывается со своим рецептором с подходящей конфигурацией [536]. И что – в любом окончании аксона сидят все эти нейромедиаторы и потенциал действия выводит в синапс полный их ассортимент? А теперь самое время познакомиться с принципом Дейла, названным по имени его первооткрывателя Генри Дейла, великого многостаночника нейробиологии. Этот принцип, предложенный в 1930-х гг., оказался настолько верен [537], что до сих пор обеспечивает каждому нейробиологу чувство глубокой внутренней правоты. Вот этот принцип: в каждом аксонном окончании нейрона потенциал действия высвобождает в синаптическое пространство один и тот же тип нейромедиатора. Поэтому у каждого нейрона будет свой характерный нейрохимический профиль. «Данный нейрон является нейроном с нейромедиатором типа А . И это означает, что нейроны, с которыми наш нейрон связан, все имеют на дендритных шипиках рецепторы нейромедиатора А » [538].
Сейчас известны десятки нейромедиаторов. Некоторые из них хорошо нам знакомы – это серотонин, норадреналин, дофамин, ацетилхолин, глутамат (самый частый возбуждающий нейромедиатор в мозге) и ГАМК (самый частый тормозный). Для студентов-медиков данная тема – настоящая пытка, им приходится выучивать все ступени синтеза [539]этих нейромедиаторов: и предшественников, и последовательность промежуточных форм, через которые проходит молекула-предшественник, пока не дойдет до финальной рабочей молекулы, да еще безобразно длинные названия ферментов, катализирующих этапы синтеза. Но все же есть некоторые относительно простые правила, построенные на трех положениях:
а) Если бежишь со всех ног от тигра, то малоприятно, когда нейроны вдруг перестают передавать сигнал мышцам, потому что – вот ведь незадача! – у них закончился нейромедиатор. Это значит, что нейромедиаторы нужно синтезировать из таких предшественников, которых в организме много; зачастую ими являются простые компоненты пищи. Например, серотонин и дофамин производятся из пищевых аминокислот – триптофана и тирозина соответственно. А ацетилхолин – из холина и лецитина.
б) Нейрон может производить в секунду десятки потенциалов действия. При каждом нужно заново заполнить пузырьки нейромедиатором, потом запустить его в синаптическую щель, а затем забрать обратно внутрь аксонного окончания. Поэтому хотелось бы, чтобы молекула нейромедиатора не была огромной, слишком сложной и с затейливыми финтифлюшками, для которых потребовались бы специальные бригады строителей. Такие молекулы, наоборот, должны синтезироваться из предшественников на раз-два, за несколько шагов. Им следует быть дешевыми и простыми в исполнении. Так, синтез дофамина из тирозина идет всего в два несложных этапа.
в) И наконец – для пущей простоты и дешевизны – синтез многих нейромедиаторов начинается с одного и того же предшественника. Так, для синтеза дофамина дофаминергические нейроны имеют два фермента, по одному на каждый этап. А в нейронах, выделяющих норадреналин, вдобавок к этим двум есть еще один фермент, он превращает дофамин в норадреналин.
Все ради удешевления. И в этом есть смысл. Ведь ничто не выходит из употребления быстрее, чем нейромедиатор, выполнивший свою постсинаптическую работу. Так вчерашняя газета годится разве что на подстилки для еще необученных щенят.
Нейрофармакология
Когда возникло понимание работы нейромедиаторов, ученые занялись вопросами, как именно действуют «нейроактивные» и «психоактивные» препараты.
Эти препараты попадают в две широкие категории: увеличивающие или снижающие возбуждение в том или ином типе синапсов. Мы уже получили некоторое представление о стратегиях повышения эффективности передачи сигнала: а) стимуляция синтеза нейромедиатора (можно для этого, например, ввести его предшественник или с помощью лекарств увеличить активность ферментов его синтеза), например, при болезни Паркинсона в одном из участков мозга перестает вырабатываться дофамин, поэтому лечение предусматривает введение вещества L – DOPA, которое является непосредственным предшественником дофамина, и в результате уровень этого гормона в мозге увеличивается; б) введение искусственно синтезированного нейромедиатора или близкого по строению аналога, обладающего достаточным сродством к рецептору конкретного нейромедиатора: так, псилоцибин структурно соответствует серотонину и активирует те же типы рецепторов; в) повышение восприимчивости постсинаптического нейрона к нейромедиатору за счет увеличения количества его рецепторов, в теории выглядит неплохо, но на практике это сделать трудно; г) увеличение количества нейромедиатора в синапсе с помощью подавления ферментов, его расщепляющих; д) продление эффекта действия нейромедиатора в синапсе путем торможения его обратного захвата. Вот, к примеру, самый ходовой антидепрессант – прозак – именно этим и занимается в серотониновых синапсах. Поэтому его часто называют СИОЗС, т. е. селективный ингибитор обратного захвата серотонина.
Между тем среди использующихся лекарственных препаратов имеются и такие, которые уменьшают уровень передачи сигнала в синапсах; механизм действия подобных препаратов легко представить. Например, они могут блокировать синтез нейромедиатора, или его выход в синаптическую щель, или работу рецепторов и т. д. Вот показательный пример. Ацетилхолин активирует сокращение диафрагмы. Яд кураре, который в ходу у племен Амазонии, блокирует рецепторы ацетилхолина. И человек перестает дышать.
И еще один важный момент: точно так же, как пороговое значение возбуждения в аксонном холмике постепенно сдвигается по мере приобретения опыта, практически вся нейромедиаторная матчасть тоже меняется.
Три и более нейрона
Мы с честью вышли к рубежу, когда можно подумать уже о трех нейронах. А через малое число страниц мы осмелеем настолько, что возьмемся рассуждать сразу о нескольких. И в этом разделе мы должны будем понять, как работает нейронная сеть, т. е. подняться на промежуточную ступень между двумя нейронами и целыми отделами мозга, заправляющими нашим поведением – самым лучшим, добродетельным и самым худшим, а то и просто ужасным.
Нейромодуляция
Рассмотрим следующую диаграмму.
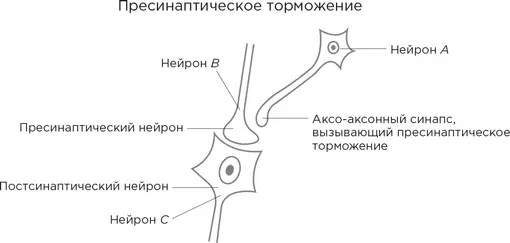
Аксонное окончание нейрона В формирует синапс с дендритным шипиком постсинаптического нейрона (назовем его нейрон С ) и выделяет в синаптическую щель возбуждающий нейромедиатор. Все как обычно. Однако есть еще нейрон А , чьи аксонные отростки идут к нейрону В . Но они заканчиваются не как у нормальных нейронов на дендритных шипиках, а на аксонном окончании нейрона В .
Читать дальшеИнтервал:
Закладка:




![Роберт Сапольски - Игры тестостерона и другие вопросы биологии поведения [litres]](/books/1074102/robert-sapolski-igry-testosterona-i-drugie-vopros.webp)




