Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Приспособленность популяций A и B (т. е. их усредненная каталитическая активность) за десять поколений не изменилась. Они не стали справляться со своей функцией лучше, чем исходный рибозим Azo . По-видимому, Azo уже был максимально оптимизирован для выполнения этой функции, и за десять поколений эволюционирующим популяциям так и не удалось «найти» такую мутацию, которая бы сделала разрезание субстрата еще более эффективным. Ни одна из возникших мутаций не оказалась полезной и не была поддержана положительным отбором. Таким образом, в течение десяти поколений действовал только очищающий отбор, который отбраковывал вредные мутации, а нейтральные игнорировал, пассивно позволяя им накапливаться.
Теперь можно было проверить главное: поможет ли скрытая изменчивость приспособиться к новым условиям. «Новые условия» состояли в том, что исследователи заменили субстрат. Вместо стандартного олигонуклеотида они предложили рибозимам его производное, в котором один из атомов кислорода в остатке фосфорной кислоты (расположенном как раз в том месте, где рибозим должен разрезать цепочку РНК) был заменен на атом серы. «Дикий» рибозим Azo справляется с таким субстратом, но с большим трудом (с низкой эффективностью).
После смены субстрата эксперимент продолжался еще восемь поколений, но теперь в нем участвовали уже не две, а три популяции. К линиям A и B добавилась третья, контрольная линия, состоящая из рибозимов «дикого типа» (WT) и не имеющая запаса скрытой изменчивости.
Все три популяции за восемь поколений в той или иной степени приспособились к новому субстрату, т. е. стали разрезать его эффективнее, чем вначале. Однако популяции A и B приспосабливались быстрее и достигли в итоге большей эффективности, чем линия WT (см. рисунок).
Секвенируя рибозимы последнего поколения, авторы обнаружили, что в популяции A наибольшей численности достиг рибозим-мутант с семью мутациями, получивший название AzoΔ . В популяции B победителем эволюционного соревнования оказался другой мутант — Azo* , у которого обнаружилось четыре мутации. Все мутации у двух «чемпионов» оказались разными.
Рибозим Azo* , как показали дальнейшие эксперименты, действительно разрезает новый субстрат с высокой эффективностью. Что же касается рибозима AzoΔ , то в чистом виде он, к удивлению ученых, вообще не смог справиться с новым субстратом. Он оказался своеобразным рибозимом-«прихлебателем»: он прекрасно режет новый субстрат только в присутствии других рибозимов (например, исходного или Azo* ). Вероятно, другие рибозимы помогают ему свернуться в правильную трехмерную конфигурацию. Подобные ситуации ранее уже были замечены в экспериментах с рибозимами [59].
Что касается линии WT, то в ней за восемь поколений так и не появились эффективные мутанты, сравнимые с Azo* или AzoΔ .
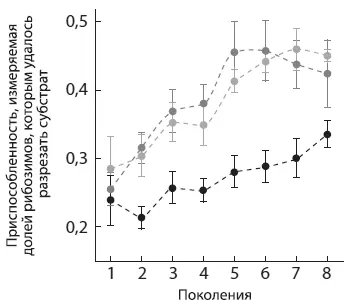
Приспособление популяций A (серые кружки) , B (светло-серые) и WT (черные) к новому субстрату. По горизонтальной оси — поколения, по вертикальной — приспособленность, измеряемая долей рибозимов, которым удалось разрезать субстрат. По рисунку из Hayden et al., 2011 .
Авторы выяснили, каким образом скрытая изменчивость помогла популяции B произвести удачливого мутанта Azo* . Оказалось, что после десяти поколений очищающего отбора в популяции уже были рибозимы с одной, двумя и даже тремя мутациями из четырех, составляющих генотип Azo* . Эти мутации по отдельности не дают преимущества ни в старых, ни в новых условиях. Для эффективного разрезания нового субстрата нужны все четыре мутации вместе. Понятно, что наличие у некоторых рибозимов в популяции B отдельных мутаций из этого комплекса резко повысило вероятность возникновения удачного генотипа Azo* . Пользуясь терминологией Ленски, можно сказать, что на стадии накопления нейтральных мутаций произошло «потенцирование» рибозимов популяции B, повысившее вероятность последующей «актуализации», т. е. закрепления ключевой мутации, создавшей новый признак.
Исследование показало полезность скрытой изменчивости для адаптации к новым условиям. Но, чтобы скрытая изменчивость могла накопиться, организмы должны быть помехоустойчивыми. Если бы исходный рибозим не обладал этим свойством, почти все мутации были бы для него в нормальных условиях вредными. Очищающий отбор безжалостно бы их отсеивал, и при смене условий в популяции не оказалось бы особей, преадаптированныхк новому субстрату.
Удивительная диалектика: пытаясь сделать организмы более устойчивыми (стабильными, неизменными), отбор в итоге повышает их эволюционную пластичность, облегчая последующие эволюционные изменения.
Эволюция в монокультуре не похожа на эволюцию в сообществе
Мы рассмотрели лишь малую часть эволюционных экспериментов, проведенных биологами в последние годы. Экспериментальное изучение эволюции сегодня продвигается семимильными шагами. В этой молодой области то, что уже сделано, — сущий пустяк по сравнению с тем, что еще предстоит сделать.
В большинстве экспериментов изучалась эволюция либо искусственных молекулярных систем, либо изолированных видов (таких опытов поставлено больше всего), либо пары взаимосвязанных видов (например, паразита и хозяина). Между тем прекрасно известно, что межвидовые взаимоотношения, коэволюция, приспособление видов друг к другу играют в развитии живейшую роль. Трудно придумать пример «чисто абиотического» фактора, на который не влияли бы (прямо или косвенно) живые организмы [60].
Присутствие других видов может повлиять на судьбу эволюционирующей популяции множеством способов. Например, из-за наличия конкурентов популяции может достаться меньше ресурсов, в результате ее численность будет ниже, поэтому придется дольше ждать появления редких полезных мутаций, усилится генетический дрейф и в целом адаптация будет идти медленнее. Или на первый план может выйти не индивидуальный, а межвидовой отбор: виды, лучше адаптированные к среде, просто-напросто вытеснят конкурентов, причем серьезные изменения в генофондах могут даже не успеть произойти. Или популяции будут искать компромисс между приспособлением к абиотическим факторам и друг к другу: в этом случае монокультуры, которым не нужно идти на такие компромиссы, должны лучше приспособиться к неживой среде, чем члены сообщества. Наконец, популяции могут научиться извлекать пользу из присутствия других видов и даже наладить с ними взаимовыгодное сотрудничество, что должно пойти на пользу сообществу как целому.
Читать дальшеИнтервал:
Закладка:








