Михаил Супотницкий - Эволюционная патология

- Название:Эволюционная патология
- Автор:
- Жанр:
- Издательство:Вузовская книга
- Год:2009
- Город:Москва
- ISBN:978-5-9502-0378-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Супотницкий - Эволюционная патология краткое содержание
Книга рассчитана на широкий круг читателей. Особенно полезной она будет для исследователей, интересующихся фундаментальными проблемами патогенеза и эпидемиологии новых инфекционных болезней, для студентов биологических и медицинских факультетов вузов, а также для врачей-инфекционистов и эпидемиологов.
Книга издана в авторской редакции.
Эволюционная патология - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Геном — полная генетическая система клетки, определяющая характер онтогенетического развития организма и наследственную передачу в ряду поколений всех его структурных и функциональных признаков. Суммарные данные о содержании разных видов последовательностей в геноме человека приведены в табл. 1.
[2] Из книги Тарантул В.З., 2003.
Тип последовательности | Содержание, %
Экзоны генов | 1
Интроны генов | 25
Транспозируемые элементы | 45
Большие дупликации | 5
Простые повторы (микросателлиты) [3] Повторяющаяся ДНК (repetitive DNA) в геноме человека включает гены, которые кодируют компоненты рибосом, организованные как тандемные повторы, повторяющиеся по 150–200 раз. Подобным же образом организована высокоповторяющаяся ДНК (highly repetitive DNA), включающая очень короткие последовательности, которые могут повторяться миллионы раз, — это так называемая минисателлитная ДНК (minisatellite DNA). Она включает блоки длиной по 10—100 пар оснований (bp), характерные для каждого индивидуума (в данной работе не рассматриваются).
| 3
Другие межгенные последовательности | 20
Классификация транспозируемых элементов. Почти половину генома человека составляют различные транспозируемые элементы (transposable elements, TEs). Они делятся на два основных класса: ДНК транспозоны (DNA transposones) и ретроэлементы (retroelements). Классификация транспозируемых элементов, их процентное содержание и приблизительное количество показаны на рис. 1.
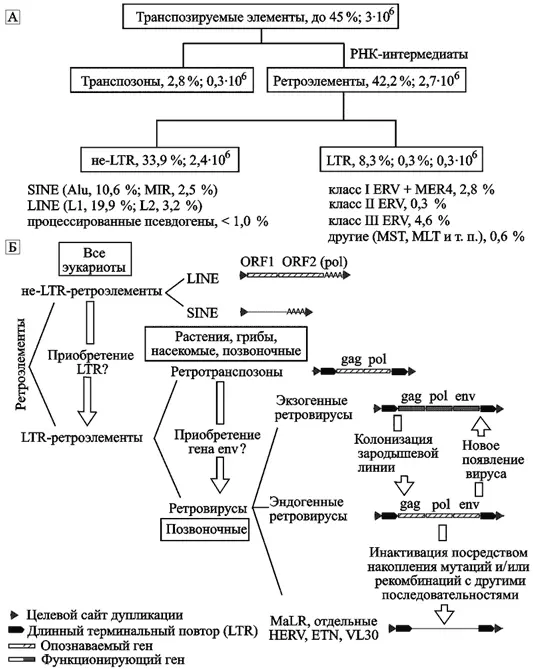
А. Общая классификация транспозируемых элементов человека по N. Bannert и R. Kurth (2004).Б. Классификация ретроэлементов по N. de Parseval и T. Heidmann (2005). Короткие прямые повторы фланкируют все ретроэлементы, интегрировавшиеся с геномом хозяина через транспозицию. ORF — открытая рамка считывания. VL30, ETN — повторяющиеся LTR-элементы, найденные у мышей. MaLR — LTR-ретротранспозон, обнаруженный у млекопитающих.
ДНК-транспозоны млекопитающих структурно сходны с бактериальными транспозонами. Подобно другим транспозируемым элементам (см. ниже), ДНК-транспозоны изменяют эволюционную траекторию своего хозяина благодаря следующим механизмам: 1) через изменение функции генов путем вставок генов, регуляторных элементов, «перетасовок» экзонов и интронов; 2) через индукцию хромосомных перестановок; 3) как источники кодирующей и не кодирующей ДНК, которая позволяет появляться различным генетическим новинкам, таким как новые гены и регуляторные последовательности (Feschotte C., Pritham E., 2007).
ДНК-транспозоны реплицируются без РНК-производного и теоретически способны перемещаться по геному по типу «разрезал и встроился» («cut and paste») через использование фермента транспозазы (transposase). Они фланкированы посредством инвертированных терминальных повторов (inverted terminal repeats, ITRs) и имеют одну открытую рамку считывания (open reading frame, ORF), которая кодирует фермент транспозазу. ДНК-транспозоны фланкированы через короткие прямые повторы (short direct repeats, DRs), «приобретенные» в ходе интеграционных процессов; не образуют вирусных частиц и не могут покинуть клетку.
Эволюционная история ДНК-транспозонов приматов закончилась еще до «расщепления» приматов на виды Старого и Нового Света. J. K. Pace II. и С. Feschotte (2007) исследовали не менее 40 семейств ДНК-транспозонов человека, включающих до 98 тыс. таких элементов, и установили высокую активность ДНК-транспозонов в эволюции млекопитающих и, в частности ранних приматов. Но перед радиацией приматов на предков видов-антропоидов (anthropoid primate ancestor), их транспозиционная активность прекратилась. Исследователи не обнаружили ДНК-транспозоны «моложе» 37 млн лет. Поэтому такие транспозируемые элементы считаются ими своего рода «окаменелостями» генома приматов (рис. 2).

А. Структура наиболее распространенного среди млекопитающих ДНК-транспозона (по Ostertag E. M., Kazazian H., 2001). Б. Суммарная активность ДНК-транспозонов в эволюции приматов. Столбики внизу рисунка соответствуют количеству ДНК-элементов, активных на отдельных этапах эволюции приматов (разбиты на суперсемейства). ДНК-транспозонов, активных после появления обезьян Нового Света, в геноме человека не обнаружено. HAT, piggyBac и Ес1/mariner — семейства ДНК-транспозонов, обнаруженные в геноме человека (по Pace II J. K., Feschotte C., 2007). В. Процентное соотношение ДНК-транспозонов и ретротранспозонов относительно общего количества транспозируемых элементов у эукариотических видов. Сокращения: Sc (Saccharomyces cerevisiae); Sp (Schizosaccharomyces pombe); Hs (Homo sapiens); Mm (Mus musculus); Os (Oryza sativa); Ce (Caenorhabditis elegans); Dm (Drosophila melanogaster); Ag (Anopheles gambiae, малярийный комар); Aa Aedes aegypti, комар, разносчик возбудителя желтой лихорадки); Eh (Entamoeba histolytica); Ei (Entamoeba invadens); Tv (Trichomonas vaginalis). (Feschotte C., Pritham E., 2007).
Млекопитающие сосуществовали с рептилиями еще в Триасе (230–190 млн лет назад), но лишь как дополнение к многообразию последних на планете. Основная масса ДНК-транспозонов (85 %; примерно 291 тыс. элементов) распространились среди млекопитающих в Меловом периоде (135-66 млн лет назад), когда происходило вымирание рептилий. «Расцвет» млекопитающих пришелся на Палеоген (66–25 млн лет назад). До 29 семейств (74 тыс. элементов) были активны у приматов перед их «расщеплением» на антропоидов, и 11 семейств (23 тыс. элементов) интегрировались с геномом антропоидов. Следовательно, в ходе эволюции приматов наблюдалось устойчивое снижение активности ДНК-транспозонов, но почему это произошло, еще предстоит установить. Более подробно о роли ДНК-транспозонов в эволюции эукариотов можно прочитать в работе C. Feschotte, E. Pritham (2007).
Ретротранспозоны . В противоположность ДНК транспозонам, они кодируют обратную транскриптазу (reverse transcriptase, RT) и перемещаются по геному через образование РНК-транскрипта. Образовавшийся транскрипт транскрибируется «обратно» в ДНК-транскрипт и встраивается в хромосому клетки. При перемещении ретротранспозонов соблюдается принцип — «копировался и вставился» («copy and paste»), как в «доброе старое время» протоклеточных образований. Ретроэлементы составляют 90 % всех транспозируемых элементов генома человека, большинство из них не активны, однако некоторые из них остались способными к ретротранспозиции.
По этим данным получается, что структуры, подобные ВИЧ, обобщенно называемые ретроэлементами, и есть геном человека. Ретротранспозируемые элементы (ретроэлементы) делят на две большие группы: способные к автономному существованию (эту их «автономность» нельзя понимать буквально, так как они зависят от ферментов репарации ДНК хозяина, необходимых для их транспозиции) и неавтономные.
Читать дальшеИнтервал:
Закладка:







![Михаил Супотницкий - Очерки истории чумы. Книга II. Чума бактериологического периода [без иллюстраций]](/books/1084186/mihail-supotnickij-ocherki-istorii-chumy-kniga-ii.webp)
![Михаил Супотницкий - Очерки истории чумы. Книга I. Чума добактериологического периода [без иллюстраций]](/books/1084187/mihail-supotnickij-ocherki-istorii-chumy-kniga-i-ch.webp)
