Михаил Супотницкий - Эволюционная патология

- Название:Эволюционная патология
- Автор:
- Жанр:
- Издательство:Вузовская книга
- Год:2009
- Город:Москва
- ISBN:978-5-9502-0378-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Супотницкий - Эволюционная патология краткое содержание
Книга рассчитана на широкий круг читателей. Особенно полезной она будет для исследователей, интересующихся фундаментальными проблемами патогенеза и эпидемиологии новых инфекционных болезней, для студентов биологических и медицинских факультетов вузов, а также для врачей-инфекционистов и эпидемиологов.
Книга издана в авторской редакции.
Эволюционная патология - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
9q34.3 | 136950603-136960065 | орангутан | 11,1-12,7 | 14
3p25 | 9864346-9871236 | орангутан | 13,4-19,8 | 14
1q23 | 163306258-163311916 | орангутан | 15,9-17,3 | 14
19p13.11B | 20248400-20258515 | орангутан | 26,4-28,1 | 14
11q12 | 61892539-61907139 | гиббон | 17,5-21,0 | 18
19q13.1 | 42289389-42298906 | гиббон | 21,0-36,3 | 18
6p22 | 28758347-28768714 | гиббон | 25,0-32,4 | 18
20q11 | 32179289-32188037 | OWM | 12,8-18,3 | 25
6p21 | 42969390-42979344 | OWM | 7,4-13,1 | 25
Процесс интеграции ретровирусов с геномом приматов не носил линейный характер. Эволюция приматов сопровождалась взрывной амплификацией ретроэлементов в их геноме. «Новые» HERV-K неоднократно занимали участки генома, принадлежащие «старым» HERV-K по механизму гомологичной рекомбинации. По крайней мере, 1/3 из исследованных провирусов подверглась эктопической рекомбинации (ectopic recombination) (Hughes J. F., Coffin J. M., 2005). J. M. Coffin (2004) указывал на периоды в эволюции приматов «без активности ретровирусов». R. Belshaw et al. (2004), исследовавшие env ген HERV восьми семейств, считают, что реинфекция является наиболее общим механизмом поддержания и пролиферации эндогенных ретровирусов в их хозяевах (см. подглаву 1.3 «Реинфекция»). Однако высокая частота этих событий затрудняет точную оценку времени эндогенизации ретровирусов на основе оценок дивергенции между LTR (рис. 5).
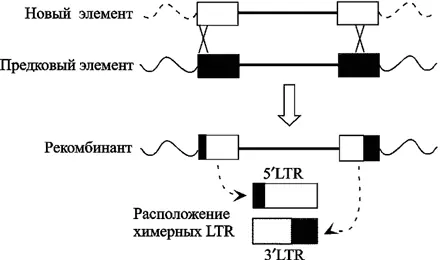
LTR заново интегрировавшихся HERV-K элементов показаны серым цветом (по Hughes J. F. и Coffin J. M., 2005).
В эволюции шимпанзе и людей участвовали разные эндогенные вирусы и с разными сценариями активности. P. Jern et al. (2006) нашли различия в недавней (т. е. имевшей место в ближайшие 5 млн лет) активности бета-подобных и гамма-подобных эндогенных ретровирусов в геномах этих видов приматов. Две большие группы гамма-подобных эндогенных ретровирусов (PtG1 и PtG2) поддерживались в геноме шимпанзе и отсутствовали у людей; PtG последовательности были наиболее сходны с двумя ERV бабуинов, но не с ретровирусами данного типа других шимпанзе или людей. Сама же гамма-ретровирусная интеграционная активность была разделена во времени от бета-ретровирусной (табл. 4).
[6] По P. Jern et al. (2006).
- | Человек | Шимпанзе |
Выявленный элемент | β | γ | β | γ |
ERV | 12 | 12 | 1 | 35
gag* [7] * интактные гены.
| 12 | 10 | 1 | 18
pro* | 12 | 4 | 1 | 27
pol* | 12 | 11 | 1 | 27
env* | 12 | 2 | 1 | 22
«LTR-gag-pro-pol-env-LTR» | 12 | 1 | 1 | 1
Для исследователей роли ретроэлементов в эволюции человека должно представлять интерес и обнаружение D. J. Hedges et al. (2004) разных сценариев эволюционной активности Alu-элементов, также начавших свой отсчет после дивергенции видов H. sapiens и P. troglodytes (см. «Эволюционная роль Alu-элементов»). Еще более любопытные результаты дает сравнительный анализ экспрессии эндогенных ретровирусов в различных тканях разных видов приматов. Например, анализ 215 образцов РНК, полученных из мозга людей, показал явную специфичность экспрессионного профиля HERV разных семейств и классов (Frank O. et al., 2005).
Недавние эксперименты позволили установить, что фундаментальные биологические различия между видами приматов являются следствием не столько вариаций в их генах, сколько результатом различий в экспрессии и регуляции одних и тех же генов (эволюция по типу анагенеза). Например, исследования, основанные на микроанализе ДНК, показывают, что экспрессия сложных генов человеческого мозга значительно превышает их же экспрессию у нечеловекообразных приматов.
Но ткани, иные чем мозг, у этих же приматов не показывают значительных различий в экспрессии генов (Stengel A. et al., 2006). А. Stengel et al. (2006) сообщили о собственных экспериментах по оценке экспрессии генов HERV в различных тканях приматов разных видов. Ими установлено, что большинство анализируемых HERV активно экспрессировались в тканях мозга человека, но оказывались либо полностью неактивными в аналогичных тканях обезьян Старого Света, либо их экспрессия была незначительной. Данные, полученные O. Frank et al. (2005) и А. Stengel et al. (2006), интересно сопоставить с более ранними наблюдениями палеоантропологов по эволюции мозга человекообразных приматов, обобщенных в работе С. Оппенгеймера (2004). Его собственные объяснения эволюции человека сводятся к необходимости приспособления приматов к внешним факторам, среди которых он на первое место ставит похолодание климата Земли, начавшееся 7–8 млн лет назад. И в качестве адаптивного признака к холоду антрополог Оппенгеймер почему-то видит увеличение объема мозга человекообразных приматов, а не увеличение длины их шерсти или толщины костей черепа. Проанализируем собранные им данные применительно к вышеуказанным работам и к результатам исследования дивергенции видов приматов, полученных другими авторами (см. рис. 4).
По данным антропологических исследований, примерно 7–8 млн лет назад произошло резкое сокращение числа видов человекообразных приматов, совпавшее по времени с расширением площади безлесых травяных степей и глобальным похолоданием, продолжавшимся несколько миллионов лет. Но именно в этот период произошла дивергенция какого-то неизвестного вида приматов на виды, в последующем дивергировавшие на гоминоидов (наших ближайших предков), горилл, орангутанов, бабуинов и шимпанзе. «Списать» оба эти процесса только на «похолодание» не удается, так как тогда же вспыхнули массовые эпизоотии ретровирусных инфекций, оставивших в качестве «отпечатков» в геноме этих видов не менее семи типов эндогенных ретровирусов. Эпизоотии были настолько масштабными, что почти не сохранили в геноме выживших видов приматов «следов» других подобных эпизоотий за предшествующие несколько миллионов лет (см. табл. 3). Массовая гибель приматов снизила заполненность занимаемых ими экологических ниш и способствовала увеличению темпов видообразования у тех представителей их отряда, которые «прошли» через процесс эндогенизации новых ретровирусов. Тогда же стали появляться виды приматов (гоминоиды), которых сегодня палеоантропологи считают нашими предшественниками, т. е. с увеличенным объемом мозга. Следовательно, ретровирусная эндогенизация при условии освобождения экологических ниш, создала условия для эволюции отряда приматов по типу кладогенеза (рис. 6).
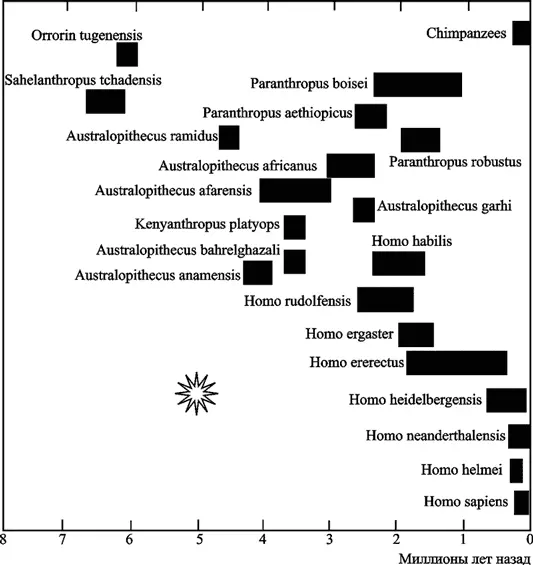
Произошла 5–2 млн лет назад после процессов эндогенизации ретровирусов, наиболее представленных в нашем геноме в настоящее время. «Звездочкой» обозначена массовая ретровирусная эпизоотия, ее «следы» сегодня обнаруживаются в геноме приматов в качестве эндогенных ретровирусов (за основу взята схема Оппенгеймера С., 2004).
Читать дальшеИнтервал:
Закладка:







![Михаил Супотницкий - Очерки истории чумы. Книга II. Чума бактериологического периода [без иллюстраций]](/books/1084186/mihail-supotnickij-ocherki-istorii-chumy-kniga-ii.webp)
![Михаил Супотницкий - Очерки истории чумы. Книга I. Чума добактериологического периода [без иллюстраций]](/books/1084187/mihail-supotnickij-ocherki-istorii-chumy-kniga-i-ch.webp)
