Михаил Супотницкий - Микроорганизмы, токсины и эпидемии

- Название:Микроорганизмы, токсины и эпидемии
- Автор:
- Жанр:
- Издательство:Вузовская книга
- Год:2006
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Супотницкий - Микроорганизмы, токсины и эпидемии краткое содержание
Кроме того, в монографии, впервые для отечественной литературы, обстоятельно освещены эпидемиология биологического террористического акта и методология выявления такого преступления. Книга хорошо иллюстрирована, содержит приложения и рассчитана на широкий круг специалистов.
Микроорганизмы, токсины и эпидемии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Ботулинические токсины связываются с рецепторами на поверхности пресинаптической мембраны двигательных нейронов периферической нервной системы и вызывают протеолиз белков в нейронах. Это приводит к ингибированию высвобождения ацетилхолина и к предотвращению мышечных сокращений — возникает вялый паралич [Halpern J., Neale E., 1995]. Столбнячный токсин сначала связывается с рецепторами на пресинаптической мембране моторных нейронов, но затем, с помощью ретроградного везикулярного транспорта, он перемещается в нейроны спинного мозга. Спастический паралич возникает из-за того, что рассечение везикуло-ассоциированных белков и синаптобревина в нейронах, нарушает высвобождение глицина и гамма-амино-битуриковой кислоты, прекращающих мышечное сокращение [Arnon S., 1997].
Активаторы иммунного ответа . Отдельные бактериальные токсины могут действовать непосредственно на Т-клетки и антигенпрезентирующие клетки иммунной системы. Самое большое семейство токсинов данного типа называют токсинами-суперантигенами (PTSAg).
Как правило, иммуностимулирующий потенциал таких токсинов является следствием их способности связывать различные участки белков главного комплекса гистосовместимости II типа, экспрессированных на поверхности антигенпрезентирующих клеток и Vбета-элементы на Т-клеточном рецепторе [Smitt С. et al., 1999]. В частности, В-домен стафилококкового TSST-1, связывает альфа-цепь антигена DR1 человеческого лейкоцита, одновременно его А-домен специфически связывается с Vбета-элементами Т-клеточного рецептора [Schlievert P., 1997]. Связывание TSST-1 с Vбета2 приводит к массивной пролиферации более 20 % периферических Т-клеток. Следствием Т-клеточной экспансии является массивное высвобождение интерлейкинов (1, 2 и 6 типов), гамма-интерферона, факторов некроза опухолей (альфа и бета) и др. [Schlievert P., 1997]. Совместно эти цитокины вызывают гипотензию, высокую температуру и диффузные эритематозные высыпания [Smitt С. et al., 1999]. Токсины данного типа характерны для случайных и факультативных паразитов.
Трехсоставные токсины . К ним мы относим структуры типа A1-B-A2, где В — это субъединица, участвующая в связывании токсина с рецептором, А1 — и А2 — субъединицы, проявляющие различную энзиматическую (токсическую) активность в клетке хозяина. Наиболее изученным токсином данного типа является сибиреязвенный. Он состоит из В-субъединицы, называемой протективным антигеном (ее используют для иммунизации против B. anthracis) и двух ферментативных субъединиц (А-субъединиц), одна из которых — отечный фактор (кальмодулинзависимая аденилатциклаза), индуцирует образование вторичных мессенджеров (цАМФ); другая — летальный фактор, является металлопротеазой (см. табл. 3).
Введение чувствительным животным любой из этих субъединиц по отдельности, не приводило ни к каким патологическим последствиям. Внутрикожное введение комбинации В-субъединицы и отечного фактора, приводило к развитию кожных отеков у морских свинок и кроликов. Смесь В-субъединицы и летального фактора при внутривенном введении вызывала гибель мышей и крыс, но не давала образования отеков при внутрикожном введении. При введении смеси трех компонентов токсина, они действовали синергически в тестах проверки летальности на мышах [Бургасов П.Н., Рожков Г.И., 1984]. В более поздних исследованиях было показано: что отечный фактор вызывает только кратковременное повышение внутриклеточного уровня цАМФ, поскольку он быстро разрушается Клеточными протеазами [Motecucco С. et al., 1994]. Т. е. развитие сибиреязвенной интоксикации предполагает обязательное участие всех трех компонентов сибиреязвенного токсина.
С. Petosa и соавт. (1997), выявили тонкие механизмы этого процесса. В-субъединица (протективный антиген — ПА, РА) сибиреязвенного токсина представляет длинную плоскую молекулу, размером 100х50х30 ангстрем, состоящую из 4 доменов. Связывание ПА с рецептором клетки-мишени начинается с домена 4. Этот домен (остатки 596–735) имеет первичную шпильку и спираль, за которыми следует бета-сендвич с иммуноглобулиновой складкой. Домены 1, 2 и 3 тесно связаны между собой, но домен 4 имеет с ними ограниченный контакт. Внутри иммуноглобулиновой складки домена 4 содержится доступная петля из 19 аминокислотных остатков, аналогичная антигенсвязывающей CDRS-петле антител и рецепторсвязывающей петле дифтерийного токсина. Протеолитическая активация происходит на клеточной поверхности. Протеаза фурин (она же используется для протеолитическои активации дифтерийного токсина, экзотоксина А псевдомонад и некоторых вирусов) расщепляет поверхностную петлю внутри домена 1. В результате высвобождается N-терминальный 20 кд фрагмент (ПА20). Этот фрагмент не играет какой-либо дополнительной роли в интоксикации. Однако его удаление приводит к образованию большой гидрофобной поверхности на оставшемся фрагменте ПА (ПАб3). Остальная часть домена 1, называемого теперь домен 1', образует N-окончание активного ПАб3. После утраты ПА20. ПАб3 формирует гептамер, который вставляется в мембраны при кислых значениях рН, формируя катионселективные каналы как в искусственных липидных бислоях, так и в клетках. Новая гидрофобная поверхность домена 1' полностью обнажается, формируя часть большого, плоского гидрофобного пятна на «вершине» гептамера. Эта поверхность обеспечивает открытый сайт для связывания фактора отека и летального фактора, которые теперь связывают ПАб3 с высоким аффинитетом (рис. 18).
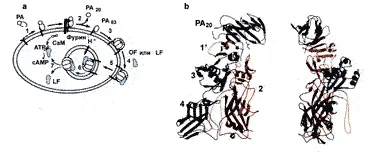
Рис. 18. Стадии сибиреязвенной интоксикации. a.1 — РА связывается с рецептором клетки хозяина; 2 — фурин расщепляет и высвобождает ПА20; 3 — ПАб3 формирует гептамер; 4 — токсические ферменты связываются с ПАбз; 5 — опосредованный рецептором эндоцитоз; 6 — подкисление эндосомы приводит к мембранной вставке ПАб3; 7 — транслокация токсических ферментов в цитозоль. LF — летальный фактор; EF — фактор отека. b. Ортогональные проекции РА, окрашенного по доменам. Домен 1 включает РА плюс домен 1' и два иона Са2+. ПА20 отщепляясь, больше не играет роли в процессе интоксикации. Образовавшаяся гидрофобная поверхность домена 1' формирует участок связывания для LF и EF [Petosaet al., 1997]
Т.е. трехкомпонентные токсины используют общую В-субъединицу, обеспечивающую ферментативным субъединицам единый механизм проникновения в цитозоль. Видимо это необходимо для проявления синергидного эффекта токсического действия ферментативных субъединиц.
Происхождение токсинов.С момента открытия Roux и Yersin в 1888 г. дифтерийного токсина, этот вопрос вызывает много споров. В последние годы появилась возможность проведения тонких молекулярных сравнений между структурами отдельных токсинов. Результаты этих сравнений оказались неожиданными для ученых [Finlay В., Falkow S., 1997]. Токсины микроорганизмов из отдаленных семейств и даже родов, занимающих различные экологические ниши и вызывающие совершенно не похожие инфекционные болезни, оказались весьма сходными по механизму действия и по критическим для проявления токсичности структурам (например, шигатоксин, коклюшный и холерный токсины). Было обнаружено сходство между токсинами и ферментами эукариотических клеток (например, отечный фактор B. anthracis и аденилатциклаза человека). По этим данным создается впечатление не только широкого эволюционного родства токсинов между собой. Но, как заметил Ю. Вертиев (1996), у некоторых исследователей даже возникает мысль, что токсины возникли путем захвата из эукариотической клетки генетического материала, кодирующего токсин. Накопление экспериментального материала по тонкой структуре и механизму действия токсинов создало условия для выдвижения интересных гипотез об их происхождении. Но, видимо, трудно рассчитывать на то, что этот вопрос будет однозначно решен для каждого бактериального токсина в рамках какой-то общей гипотезы.
Читать дальшеИнтервал:
Закладка:





![Михаил Супотницкий - Очерки истории чумы. Книга II. Чума бактериологического периода [без иллюстраций]](/books/1084186/mihail-supotnickij-ocherki-istorii-chumy-kniga-ii.webp)
![Михаил Супотницкий - Очерки истории чумы. Книга I. Чума добактериологического периода [без иллюстраций]](/books/1084187/mihail-supotnickij-ocherki-istorii-chumy-kniga-i-ch.webp)


