К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Когда Хебб писал свою книгу, существовали две основные точки зрения на функционирование нервной системы. Одна из них была хорошо обоснована, другая – нет. Первая касалась вопросов генерации нервных импульсов и их передачи через «инапсы. Другая имела отношение к электрическим полям, создаваемым потенциалами мозга. В. Кёлер построил свою гешталь-тистскую концепцию на основе этих неироэлектрических полей и потерпел неудачу, доказывая их повсеместное существование десять лет спустя после опубликования Хеббом своих взглядов.
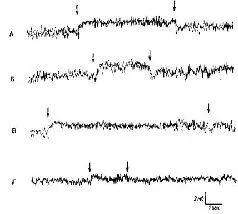
Рис. VI-8. Сдвиг постоянного потенциала (ПП) в слуховой коре в ответ на звуковую стимуляцию. «Активный электрод» – на твердой оболочке мозга над средней эктосильвиевой извилиной; другой электрод – на фронтальной надкостнице. Отклонение вверх означает негативность на твердой оболочке относительно кости. Л – сдвиг ПП в ответ на белый шум; Б – сдвиг ПП в ответ на тон 4000 гц; В – сдвиг ПП, возникающий в ответ на белый шум, возвращается к нулевой линии до прекращения звуковой стимуляции; Г – ответ на звуковые щелчки (50 в сек) и его возвращение к нулевой линии до прекращения стимуляции (Gumnit, 1960).
Я был счастлив, что смог принять участие в этих исследованиях. Экспериментальное наступление оказалось в целом успешным (КбЫег, 1958), и исследователи в моей лаборатории, а также в других лабораториях, регистрируя и вызывая в мозгу электрические поля постоянного тока, установили -их корреляции с функциями мозга и поведением (Gumnit, 1961; Stamm, 1961; Morrell, 1961). Всякий раз при стимуляции организма через тот или другой сенсорный вход (рис. VI-8) в соответствующих ограничейных областях мозга возникал сдвиг постоянного потенциала. Прямое раздражение мозга постоянным током могло задержать или ускорить научение в зависимости от полярности прикладываемого потенциала. Но я не согласился с Кёлером, когда он стал настаивать на связи между электрическими полями постоянного тока и восприятием. Позднее, когда я закончил эксперименты, в которых я накладывал на поверхность коры мозга алюминиевую пасту, мы снова смогли прийти с ним к согласию. Эксперименты показали, что различение структуры сигналов не нарушается, не-смотря на заметные изменения постоянного потенциала и ЭЭГ (рис. VI-9, VI-10).
А Б
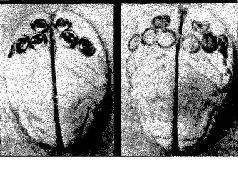
Рис. VI- 9. A – фотография мозга обезьяны со вживленными з него серебряными дисками, содержащими пасту; Б – фотография того же самого мозга после снятия дисков. В данном случав вживлепие было произведено в теменную кору. Другие эксперименты были проведены на затылочной, височной и лобной коре тем же способом или методом множественных точечных инъекций в кору алюминиевой пасты (Stamm and Warren, 1961).
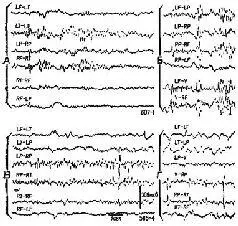
Рис. VI-10. Образец записи ЭЭГ у двух обезьян со вживленными дисками, как показано на рис. VI-9. А и Б – участки записи ЭЭГ до операции, В и Г – спустя 5 месяцев после вживления дисков с алюминиевой пастой. Биполярное отведение: L – левое, R – правое полушарие, F – лобная кора, Т – височная, Р – задняя темен-лая, V – макушка. Калибровка, как показано (Stamm and Warren. 1961).
Кёлер никогда не признавал экспериментов, проведенных Лешли (Lashley, Chow and Semmes, 1951), в которых для опровержения его теории и разрушения нейроэлектрических полей использовалась золотая фольга. Не признавал он также и экспериментов Сперри с перекрестом нервных волокон (Sperry, Miner and Meyers, 1955), в которых использовались изолирующие полоски слюды. Но когда он познакомился с результатами эксперимента со вживлением дисков с алюминиевой пастой, он воскликнул: «Это опровергает не только мою теорию поля постоянного тока, но и всякую другую современную неврологическую теорию восприятия!»
Позвольте мне коротко остановиться на тех данных, которые накопились в ходе этой дискуссии, чтобы рассеять такой мрачный взгляд на неврологию восприятия. Как было подробно показано в первой части книги, нервный импульс не является единственной важной электрической характеристикой нервной ткани. Другой характеристикой является микроструктура медленных потенциалов. Хотя последние близки к кёлеровским полям постоянного тока, они существенно отличаются от них тем, что они не диффузны, а строго локализованы на соединениях между нейронами или на дендритах и могут даже иметь вид миниатюрных спайков, которые чаще всего при распространении ослабевают. Проведение нервного импульса в любом месте нервной системы ведет к образованию на соединениях микроструктуры медленных потенциалов. Когда нервные импульсы достигают синапсов, генерируются постсинаптические потенциалы. Они никогда не бывают единичными, а образуют динамические структуры афферентных воздействий. Когда постсинаптические потенциалы возникают в дендритных полях мозга, они часто оказываются недостаточно большими, чтобы немедленно вызвать разряд нервного импульса. Поэтому динамические структуры постсинаптических потенциалов образуют узор, который имеет сходство с фронтом волны. Но этот рисунок медленных потенциалов не является каким-то таинственным фоном, миражем, наложенным на известную нервную функцию. Он представляет собой микроструктуру, состоящую из классических нейронных медленных потенциалов, возникающих в результате прихода нервных импульсов, ожидающих выхода через аксон.
Таким образом, структуры, образуемые на входе и выходе и принявшие форму микроструктуры медленных потенциалов, становятся источником третьей силы в нервной клетке. Все это говорит против концепции «плавающего» поля как возможного нейрофизиологического механизма, обусловливающего целостный характер восприятия. Потребность в этом третьем источнике была обнаружена до его открытия. Лешли глубоко волновала следующая проблема:
«Здесь мы сталкиваемся с такой дилеммой. Нервные импульсы проводятся по определенным, строго ограниченным путям, по сенсорным и моторным нервам и в центральной нервной системе от клетки к клетке и через-определенные межклеточные связи. Однако кажется, что все поведение детерминировано массой возбуждения, формой, отношением или соотношением возбуждения внутри общих полей активности безотносительно к отдельным нервным клеткам. Важна динамическая структура возбуждения, а не отдельный элемент. Какой тип нервной организации способен отвечать на такую структуру возбуждения при отсутствии специфических, ограниченных путей, обеспечивающих его проведение? Эта проблема имеет отношение к деятельности всей нервной системы, и необходимо сформулировать какую-нибудь гипотезу, чтобы дать направление дальнейшему исследованию» (Lashley, 1942, стр. 306).
Читать дальшеИнтервал:
Закладка:








