К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
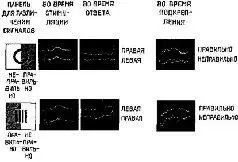
Рис VII-4 Результаты эксперимента на зрительное различение у обезьяны, условия его показаны на рис VII-3 Для характеристики реакции биотоков мозга на действие стимула в течение 3 дней производилось усреднение участков ЭЭГ, регистрируемых непосредственно после появления на панели круга или полос Для характеристики ответов животного использовалась запись ЭЭГ, непосредственно предшествующая тому моменту, когда обезьяна отвечала на стимул нажатием либо на левую, либо на правую половину панели Запись ЭЭГ производилась также во время подкрепления (арахисом), если обезьяна правильно отвечала на стимул, и в отсутствие подкрепления, еои она ошибалась
То, что видит обезьяна – полосы или круг, – находит отражение в различии волновых форм (второй впадины W – волны), возникающих на стимулы Только после того, как она научается правильному выпот-нению этого задания, появляются различия в волновых формах ЭЭГ, которые непосредственно предшествуют моменту нажатия левой ити правой половины панели Эти различия, по-видимому, не зависят ог того, видит ли обезьяна круг или полосы Таким образом, вотаовые формы в ЭЭГ отражают ее намерение нажать на определенную половину панели и ничего не говорят о том, будет ли ее ответ правильным или ошибочным Однако имеется третий вид различия в волновых формах, записываемых во время подкрепления Оно показывает, было ли полечено подкрепление медленный сдвиг нулевой линии, следующий за двигательной реакцией, указывает, что животное ожидает награды, а появление в ЭЭГ волн частотой 25-30 гц свидетельствует оегоразоча ровании (Pribram, 1969)
Однако другая группа экспериментов, проведенных в моей лаборатории (Pribram, Spinelli and Kamback, 1967, рис. VII-З, VII-4), показывает, что если научение уже имело место, то информация в мозгу не распределяется по всей системе. Были использованы очень маленькие электроды. Обезьяны были обучены по-разному реагировать на различные геометрические стимулы. В отличие от экспериментов Джона стимулы освещались очень короткой вспышкой света В зрительной коре было обнаружено несколько различных типов волновых форм электрической активности. Один тип был получен, когда форма волны определялась относительно момента предъявления стимула. Этот тип вызванного ответа обнаружил четкую зависимость от характеристики стимула. Два других типа были получены, когда форма волны определялась по отношению к началу ответа животного. Один тип волновой формы зависел от того, получила обезьяна подкрепляющий пищевой шарик за правильный ответ или не получила, когда ее ответ был ошибочным Другой тип волновой формы возникал непосредственно перед двигательной реакцией животного. Эта форма волны коррелировала с определенным типом двигательного ответа (нажатием на правую или левую панель); она следовала за стимулом, но не зависела от него и от полученного подкрепления. Важным для нас является тот факт, что все эти типы характерных волновых форм появляются локально в зрительной коре. Один тип волновой формы записан от одних электродов, другой от других. Распределение этих волновых форм происходило не в соответствии с какой-либо структурой возбуждения. Однако со дня на день, от недели к неделе наблюдалось полное постоянство в записях, получаемых от каждого отдельного электрода. Какой бы процесс кодирования ни происходил, он стабилизировался по мере того, как проводилась запись.
Результаты этих экспериментов не согласуются с представлением о том, что образ, подобно фотографии, проецируется на поверхность коры. Они показывают, что каждая сенсорная система функционирует, имея большой резерв. Поскольку системы с разрушением и без него обнаруживают лишь незначительные различия в поведении, этот резерв, должно быть, распределен по всей системе – сохраняемая в ней информация, необходимая для осуществления различения, дублируется одновременно многими ее участками. Таким образом, вполне вероятно, что задержка в научении, возникающая при наличии эпилептогенных очагов, вызванных наложением на поверхность мозга алюминиевой пасты, обусловлена помехами, которые вносятся этими очагами в процесс дублирования хранения информации (рис. VII-5).
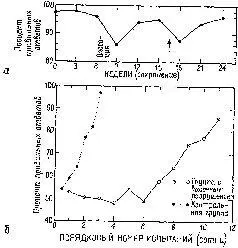
Рис. VII-5. а – сохранение зрительного различения, которое проверялось каждые 3 недели в экспериментах, аналогичных тем, которые были показаны на рис. VI-9 и VI-10. Вживление диска с пастой алюминия отмечено словом «операция»; появление первых эпилептоидиых разрядов указано стрелкой; 6 – кривые научения, полученные при выполнении задания на различение зрительных структур (каждая точка – медиана данных группы животных и каждой сотни испытаний). Белые кружки показывают выполнение задания ооезьянами с эпилептогенными разрушениями в коре височной доли, черные кружки – выполнение задания контрольной группой ооезьян (Stamm and Pribram, 1961).
Вопросы, поднятые этими наблюдениями, следует сопоставить с другим рядом проблем: каким образом предметы кажутся нам достаточно стабильными, так что мы можем узнавать их независимо от угла зрения и расстояния до предмета? Каким образом мы узнаем предмет, несмотря на то, что светом, идущим от предмета, возбуждается только часть сетчатки, а следовательно, и мозга? Константность восприятия предметов и величины существует у детей уже в возрасте нескольких недель. Таким образом, простое объяснение феномена константности восприятия в терминах научения ставится под сомнение. Какого же рода механизм мог бы обеспечить одновременно существующую гибкость восприятия и константность узнавания?
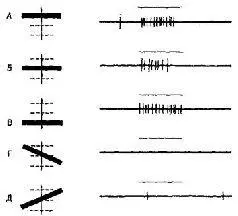
Рис. VII-6. Кортикальная клетка активируется только при освещении определенной области левого (противоположного) глаза. Эта область имеет размеры примерно 5 X 5° и расположена на 10 выше и левее центра. Нейрон лучше всего отвечает на предъявление черного горизонтального прямоугольника, который может быть без изменения эффекта параллельно смещен в пределах рецептивного поля на 1/3 X 6° (А- В). Наклонное положение прямоугольника делает его неэффективным (Г – Д). Отметка предъявления стимула показана горизонтальной линией над каждой записью нейронной активности (Hubel and Wiesel, 196).
Как факты восприятия структур при наличии скотомы, так и факты константности восприятия говорят о том, что существует, должно быть, какой-то эффективный нейрологический механизм пространственного распределения информации на входе мозга. Для объяснения фактов восприятия недостаточно изоморфизма, простого («точка в точку») соответствия между стимулом и картиной возбуждения в коре, обусловленной анатомией системы. Когда в результате удаления коры происходит выпадение 80% зрительного поля, узнавание осуществляется благодаря оставшейся части зрительного поля; когда в зрительной коре производится множество разрушений, то сохранившиеся части функционируют столь успешно, что при различении испыты-ваются лишь незначительные трудности; рассматриваем ли мы предмет той или другой частью сетчатки, под тем или иным углом зрения, мы всегда можем узнать его. Восприятие отличается от обычного фотографического изображения – оторвите 98 или даже 80% У фотографии и попытайтесь идентифицировать ее: в большинстве случаев это невозможно.
Читать дальшеИнтервал:
Закладка:








