К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Один из способов решить эту дилемму – это отказаться от представления о том, что нейронный механизм входа является в каком-либо отношении устройством, формирующим образ, предположить, что узнавание структур есть результат операции выделения признаков, имеющей место на самом входе и возникающей с помощью отдельных нейронов или их небольших групп, и что восприятие зрительных структур осуществляется посредством иерархической системы таких «детекторов признаков». В пользу такого представления говорит то, что внутри частей входных каналов происходит конвергенция сигналов.
Можно получить карты зрительных рецептивных полей для нейронов, находящихся на разных уровнях входной системы.
В коре мозга было обнаружено два типа таких нейронов. Примерно у половины из них эти карты имеют форму более или менее округлых полей, аналогичных рецептивным полям ганг-лиозных клеток сетчатки, но в отличие от последних не имеющие четко выраженной окружающей зоны (Jung, 1961; Spinelli, Pribram and Bridgeman, 1971). Другая половина кортикальных нейронов со значительно большей вероятностью активируется входными системами, имеющими не округлые, а удлиненные формы. Чтобы вызвать максимальный эффект, эти удлиненные формы должны быть определенным образом ориентированы в отношении глаза, на который они воздействуют; разные нейроны отвечают на различную длину и ориентацию линий, границ и даже углов (Hubel and Wiesel, 1962; рис. VII-6).
Известное объяснение этих наблюдений состоит в том, что нейроны, чувствительные к воздействию линии или границы, активируются только тогда, когда возбуждается большая часть или все линейное множество нейронов, чувствительных к воздействию таких пространственных признаков (рис. VII-7) и связанных с рецепторами линии и края. Высказано предположение, что благодаря дальнейшей конвергенции активность детекторов линии и края может комбинироваться и создавать клетки, реагирующие на более сложные структуры. Это предположение подтверждается тем, что найдены нейроны, особенно чувствительные к таким сложным структурам, как углы и короткие участки линий.
Таким образом, объяснение восприятия на основе механизма анализа признаков становится крайне соблазнительным. Электрофизиологический анализ рецептивных полей клеток внутри вход-пых каналов показывает, что выделение признака в какой-то степени происходит внутри этих каналов. Далее, есть основание предполагать, что выделение основных признаков, таких, как цвет (De Valois, 1960; De Valois and Jacobs, 1968), контур и направление, осуществляется врожденными, встроенными в организм детекторами. Этот тип механизма, осуществляющего детекцию основных признаков, дополняется механизмом, обеспечивающим выбор признака, который зависит от прошлого опыта организма. Возникает вопрос, изменяет ли прошлый опыт сами эти детекторы признаков или в результате этого опыта в деятельность вовлекаются дополнительные нейроны, вызывающие изменения в нейронной популяции, изучаемой экспериментатором. Однако очевидно, что в зрелом организме процесс выделения признаков не может быть ограничен работой встроенных в организм детекторов. Должен происходить также анализ признаков посредством нейронов памяти. Поэтому обратимся теперь к логике той архитектоники нейронов, которая присуща анализаторам признаков, чтобы определить, что можно и что нельзя объяснить с этих позиций в самом восприятии.
Софистическое утверждение Г. Вернера (1971), основанное отчасти на его собственных обширных исследованиях и работах В. Маунткасла (1957), Д. Хьюбела и Т. Визела и отчасти на концепции Т-О-Т-Е, относится к процессу анализа признаков в соместетической чувствительности и может быть с небольшими изменениями отнесено также и к другим входным системам. В коре мозга он различает базальную структуру – колонку, каждый нейрон которой обладает рецептивным полем, образующим единицу представительства входа. Эти нейроны имеют, как правило, идентичные или по крайней мере сходные рецептивные поля и, таким образом, образуют одинаковый уровень представительства. Эти колонки в свою очередь образуют более сложные структуры благодаря нейронам с дирекционпой чувствительностью, которые выполняют функцию указателей для согласования деятельности колонок (рис. VII-7).
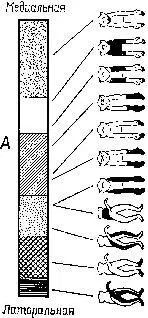
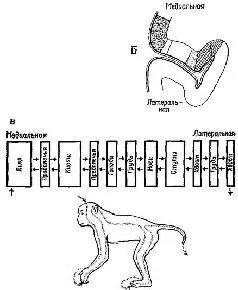
Рис. VII-7. Схематическое представление организации соматосенсорной коры в терминах рецептивных полей отдельных клрток. А – экспериментальные данные, Б – обобщение результатов, касающихся кортикальных колонок, полученных в нескольких микроэлрктродных проходках у одной, или более обезьян; В – представление об организации коры, основанное на совокупности всех данных, в терминах концепции Т – О – Т – Е, демонстрирующее проекционное отношение между организацией коры и поверхностью тела. Структура коры сравнима с кодированием информации при последовательном изложении инструкции в форме единиц Т – О – Т – Е, из которых составляются программы для компьютеров. Ср с рис. VII-8 (см. Werner, 1970).
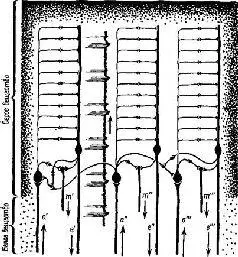
Рис. VII-8 ОККАМ Спинелли. Объяснение на стр. 148, а – входные нейроны: е – выходные нейроны, т – тестирующие нейроны. Ср. с рис. VII-9 (Pribram, 19696).
Эти указатели в зависимости от того, на какой ответ они настроены, определяют тот или иной тип взаимоотношений в электрической активности колонок; если имеются нейроны-указатели, которые чувствительны более чем к одной ориентации сигнала, то группы колонок оказываются связанными таким образом, что формируют «кольцевые структуры». Вернер сравнивает такую структуру коры с последовательным изложением инструкций, из которых составляются программы для электронно-вычислительных машин. Каждый лист программы содержит пункты, которые указывают на переходы к другим листам. Таким образом, посредством такого простого устройства могут быть запрограммированы сложные взаимодействия, последовательность операций. В терминах концепции Т-О-Т-Е наличие у колонок коры рецептивных полей со статическими свойствами создает условия для осуществления фазы проб в работе сервомеханизма; свойства колонок, свидетельствующие об их чувствительности к направлениям, составляют основу операционной фазы и фазы выхода. Сшшелли (1970) фактически составил программу (названную «Оккам»), моделирующую работу анализатора признаков, построенного па оспове такой кортикальной структуры. Предъявляя структуры нервных импульсов или волновых форм, можно настроить эту программу таким образом, чтобы она реагировала при повторении волн определенной формы (рис. VII-8). Несколько упрощепно точка зрения Вернера и Спинелли на анализатор признаков заключается в следующем: колонка коры состоит из входных и операционных нейронов, интернейронов и тестовых клеток. Входной сигнал, доходящий до нейронной единицы колонки, которая имеет свое рецептивное поле, распределяется по интернейронам. Последние в свою очередь связаны с операционными нейронами. Интерпейроны способны к настройке, то есть они адаптируются, обнаруживают привыкание и обладают памятью. Таким образом, каждый интернейрон действует как сумматор в электронно-вычислительной машине, который усредняет динамические структуры, вызванные поступающими на вход сигналами. Только при повторении таких динамических структур возбуждения в результате суммации возникает устойчивая структура, неповторяющиеся динамические структуры просто повышают нулевую линию и среднюю величину выходного сигнала. Таким образом, операционный нейрон, чувствительный только к динамическим структурам возбуждения, активируется лишь при повторении структуры входных сигналов. В целом процесс достигает своей завершенности благодаря посылке выходного сигнала от операционного нейрона к входному через тестирующий нейрон, который сличает структуру нейронной активности, вызванную входным сигналом, со структурой возбуждения, возникающей в операционных нейронах. Когда обе динамические структуры совпадают, тестирующая клетка продуцирует выходной сигнал, в противном случае при их несовпадении продолжается процесс настройки. Таким образом, каждая кортикальная колонка создает энгр:шму в соответствии со своей специфической чувствительностью к данной структуре нейронной активности, которая выступает в виде «описания» либо последовательности межспайковых интервалов, либо в волновой форме, которая являет собой огибающую узора спайковой нейронной активности.
Читать дальшеИнтервал:
Закладка:








