К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Подобные карты были пелучены для кожной чувствительности. По этому же принципу устроен и кохлеарно-кортикальный слуховой механизм (Rose and Woolsey, 1949).
Такая рецепторно-кортикальная организация не сопровождается прямым однозначным соответствием посредством единой нервной нити. Скорее всего, столь же сложная организация, которую мы уже анализировали на уровне сетчатки, повторяется как на каждом уровне механизма рецепторной системы, так и в коре. Фактически наблюдается общее сокращение числа клеток в ганглиозном слое сетчатки по сравнению с рецепторным (у человека это уменьшение оценивается примерно как 100 или 50 к одному). При переходе от слоя ганглиозных клеток к латеральному коленчатому телу таламуса число зрительных элементов, по существу, остается постоянным. При переходе от таламуса к коре возникает обратная картина: одиночная .клетка латерального коленчатого тела может контактировать с 5000 кортикальных нейронов каждый из которых находится в контакте еще примерно с 4000 других нейронов через их дендритные связи. Благодаря такому механизму тормозных взаимодействий при стимуляции у обезьяны двух точек сетчатки в фовеа, несмотря на некоторое перекрытие зон возбуждения, возникает их раздельное восприятие, причем таким образом, что двум угловым минутам на сетчатке соответствует расстояние в 1 мм на поверхности коры (Talbot and Mar-chall, 1941, p. 134). Казалось бы, что такое строение нервной системы обеспечивает проекцию некоторого «образа» от рецеп-торпой поверхности на поверхность коры во многом так же, как проецируется изображение в кинокамере. Парадокс проявляется, когда в результате болезни или хирургической операции повреждаются системы входа. Правда, как и ожидалось, в некоторых случаях может быть обнаружено выпадение части поля зрения (скотома) в месте, которое может быть предсказано в соответствии с анатомической организацией мозга (рис. VII-2).
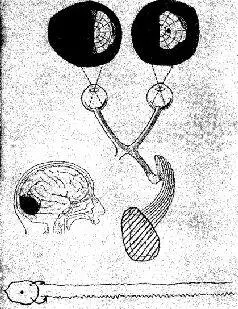
Рис VII-2. Карта зрительного поля, анатомическая схема зрительной системы 20-летней девушки с разрушением в правой затылочной долей ЭЭГ обеих зрительных долей (Case, 1942).
Однако при сохранении хотя бы малой части входного механизма это выпадение часто не замечается даже в том случае, если глаза удерживаются в неподвижности. При этом распознавание структуры сохраняется и во многих отношениях не отличается от нормального. Люди с громадными скотомами либо совсем не знают о них, либо могут быстро научиться легко их игнорировать. Животное, у которого было удалено или разрушено от 80 до 98% нервных связей входного механизма, способно решать задачи, требующие различения структур, отличающихся только деталями. Лешли (1929) удалял у крыс 80-90% стриарной коры, не нарушая их способность различать зрительные структуры. Р. Галамбос перерезал у кошек 98% волокон зрительного тракта, и животные еще могли хорошо выполнять задания, требовавшие различения весьма сходных фигур (Galam-bos, Norton and Frommer, 1967). Недавно Чау (1970), также экспериментируя с кошками, разъединял более 3/4 волокон зрительного тракта и удалял более 3/4 зрительной коры; в таких условиях едва ли оставалась неповрежденной любая из проекционных систем, характеризующихся точечным («точка в точку») представительством. Хотя зрительное различение структур после такого значительного разрушения вначале оказалось нарушенным, животное снова научилось выполнять задание, на что ему потребовалось примерно такое же число проб, как и до операции.
В моих экспериментах, как связанных с клинической нейрохирургией, так и проведенных в лаборатории (например, Wilson and Mishkin, 1959), удаление мозговой ткани было локальным и ограничивалось корой, так что оно не вызывало массивного разрушения белого вещества, благодаря чему у пациента или подопытного животного сохранялась в значительной мере способность к восприятию. После временно возникающей скотомы, длящейся несколько недель, можно было обнаружить лишь очень незначительные нарушения зрительной функции.
Как уже отмечалось, исследователи пытались использовать и многие другие методы для разрушения предполагаемой ими организации входных систем мозга, но безуспешно: Р. Сперри и его группа (1955) хирургически производили перекрестное рассечение области, воспринимающей сенсорные сигналы, и даже помещали полоски слюды в места рассечения, чтобы электрически изолировать маленькие квадратики ткани друг от друга. Лешли, Чау и Семмез (1951) пытались накоротко замкнуть электрическую активность мозга, накладывая полоски золотой фольги на воспринимающие поверхности. Я создавал в воспринимающей области коры множество эпилептогенных очагов посредством инъекции микроскопического количества алюминиевой пасты (Kraft, Obrist and Pribram, 1960; Stamm and Pribram, 1961; Stamm and Warren, 1961). Хотя такое множество очагов заметно задерживало научение различению зрительных структур, оно не препятствовало его проявлению, если научение уже закончилось или происходило после того, как были произведены многочисленные разрушения. Эти результаты показывают, что эффект входного сенсорного воздействия на мозговую ткань, получаемая на входе информация распределяются по всей входной системе.
Запись электрической активности мозга также свидетельствует о том, что в мозгу происходит распределение информации. Э. Р. Джон (John, Herrington and Sutton, 1967), например, при изучении зрительной системы использовал технику «меченых стимулов», предъявляя кошкам стимулы, которые различались не только своей геометрической структурой, но и частотой образующих эти стимулы мельканий. Эта различная частота мельканий отражалась на электрической активности мозга в ответ на предъявление стимула (или, если это мелькание было очень быстрым, то возникала субгармоника этой частоты). Таким образом, кодируемое различие частоты может оставить «след» в мозгу. Эта техника дала ряд интересных результатов, но для нас здесь важнее всего то, что тщательный анализ меченых волновых форм (вычисление позволяет обнаружить различие в волновых формах, возникающих в различных участках мозга) показывает, что одинаковые меченые волновые формы возникают во многих структурах мозга одновременно.
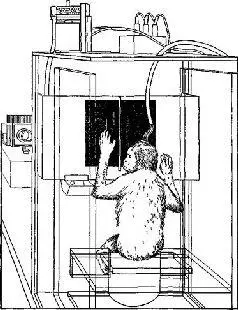
Рис VII-3 Рисунок, показывающий выполнение обезьяной задания на зрительное различение На находящейся перед обезьяной полупрозрачной панели она видит либо круг, либо вертикальные почосы, которые освещаются вспышками света в течение микросекунды Если обезьяна нажимала на правую половину панели при появлении круга и на левую – при появлении полос, то она шпучала вознаграждение – арахис, который падал в ящик под ее левой рукой Когда у нее вырабатывался навык в выполнении этого задания, электроды в зрительной коре обезьяны регистриро-1али волны определенной формы в ответ на структуру световою сигнала В начале экспериментов форма волн показывает, видит ли обезьяна круг или полосы По существу, они заранее сообщают, на какую половину панели будет нажимать обезьяна и будет ли ответ правильным или ошибочным (Pribram, 1969).
Читать дальшеИнтервал:
Закладка:








