К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Само понятие «уровень» у сторонников теории активации, судя по их анализу, весьма неопределенно. Уровень активации – это не какое-то количественное изменение интенсивности, энергетического уровня центральной нервной системы, а нарушение равновесия, организации, измеряемой количеством неопределенности (и, следовательно, количеством информации), которое характеризует системы. На нейронном уровне активация сопровождается изменением динамических структур возбуждения в ретикулярной формации ствола мозга (см. Huttenlocher, 1961) и в коре (Burns, 1968, гл. 5), а не общего количества спайковых разрядов нейронов. Даже первоначальное определение активации как электроэнцефалографической реакции основано на том наблюдении, что в ЭЭГ возникает низковольтная высокочастотная активность и что такая активность указывает на десинхронизацию. Как было отмечено Линдсли (1961), это предполагает, что во время десинхронизации нервные элементы становятся функционально независимыми друг от друга и, таким образом, способными функционировать в качестве отдельных каналов, проводящих различную информацию. Эту возросшую сепарацию можно описать как увеличение степени организации или в терминах теории информации как увеличение неопределенности (рис. XI-5).
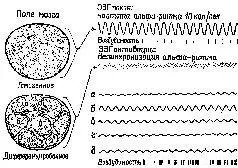
Рис. XI-5. Схема гипотетических полей мозга и связанных с ними типов ЭЭГ. Альфа-ритм покоя на частоте 10/сек характеризует состояние гомогенности, или релаксации, мозга. Десинхронизированная, активированная ЭЭГ связана с переходом к дифференцированной структуре динамических полей мозга. Активированная ЭЭГ характерна для ситуации, требующей внимания и решения задач и вообще является характеристикой более эффективного восприятия и успешной деятельности. Исходя из того, что цикл возбудимости мозга связан с подъемом и падением амплитуды волн ЭЭГ, очевидно, что при ЭЭГ покоя сменяющие друг друга периоды возбудимости и невозбудимости могли бы возникать только с частотой 10 раз в сек. В случае же десинхронизированной ЭЭГ, возникающей в результате дифференциации мозгового ноля, периоды возбудимости следуют почти непрерывно (Linds1еу, 1961).
Естественным следствием такого понимания было бы предположение, что активация представляет собой состояние, при котором независимая активность нейронных ансамблей создает свободу для «резонанса» с другими, которые, несмотря на пространственную удаленность, имеют с ними общие характеристики. Таким образом, рассредоточенная информация в состоянии активации легче направляется по нужному адресу.
Гипотеза о том, что активация является в какой-то мере механизмом неопределенности, может показаться нам привлекательной. Но это еще не все, что известно в отношении активации. Например, в эксперименте, направленном на изучение того, какое влияние в ранний период жизни оказывает ограничение движений мы обнаружили, что инактивация младенца сопровождается увеличением лабильности автономной нервной системы (Konrad and Bagshaw, 1970).
Но прежде чем будут выяснены отношения между неопределенностью и контролем за поведением, нам необходимо еще раз вернуться к теории сервомеханизмов.
Результаты Шехтера еще не получили своего объяснения: усиления того или другого переживания в результате инъекции адреналина можно достигать путем количественного изменения состояния неопределенности, вызываемого введением определенного агента; специфичность же таких чувств, как враждебность или дружелюбие и т. д., еще требует объяснения.
Специфичность чувств предполагает, что они являются скорее мотивами и эмоциями, чем просто количественным нарушением равновесия и неопределенностью или только нарушением протекания психических процессов. И конечно, в их основе лежит работа более сложной системы, чем гипоталамический механизм. Разнообразие переживаний интереса свидетельствует о том, что в устранении нарушений равновесия, в установлении специфических типов контроля, имеющих дело со специфическими ожида ниями (см. Zimbardo, 1969), участвует множество различных процессов.
Единственная возможность у организма управлять собой – это что-то делать, воздействовать на свое окружение. Как будет подробно описано в гл. XVI, всякий раз, когда сервосистема приобретает стабильность, появляются новые чувствительные индикаторы ее состояний и вводятся новые средства для управления ими. Например, когда термостаты были впервые установлены в домах, жители вначале столкнулись с более точными показателями снижения температуры: остывание наружных стен с закатом солнца вызывало поглощение тепла этими стенами. Поэтому были установлены термостаты для наружных стен, которые обеспечили более многосторонний контроль за отоплением в домах. Тот факт, что механизмы контроля развиваются как бы по спирали, обычно упускается из вида в самых распространенных определениях биологического гомеостатического процесса, что дает основание для возражений, выдвигаемых против биологической теории сервомеханизмов.
Но организм может достигать различных форм контроля не только посредством действия. Он может управлять, осуществляя самоконтроль, то есть он может создавать на основе своей неврологической системы внутренние регуляторы, – регуляторы, которые будут приводить к восстановлению равновесия даже и без внешнего действия. Положение, из которого мы исходим, заключается в том, что эти внутренние механизмы приспособления являются тем, что мы переживаем как эмоции.
Сейчас мы располагаем надежными нейрофизиологическими данными о том, что такие внутренние регуляторы не только возможны, но что указание на них стало уже общим местом. Было проведено множество экспериментов, чтобы показать, что входные каналы организма и даже сами рецепторы являются объектом контроля со стороны центральной нервной системы. Серии экспериментов, недавно выполненные в моих лабораториях, показали наличие кортикофугальных (кортико-подкорковых) влияний так называемых ассоциативных областей на периферию: на кохлеарное ядро и зрительный тракт (Dew-son, Nobel and Pribram, 1966; Nobel and Dewson, 1966; Spinelli and Pribram, 1966, 1967; Spinelli, Pribram and Weingarten, 1965; Spinelli and Weingarten, 1966; Weingarten and Spinelli, 1966). Было показано, что при стимуляции коры на этих уровнях нервной системы изменяется электрическая активность, вызванная щелчком и вспышкой света; в этих случаях изменяется как величина, так и форма рецептивных полей нейронов зрительной системы (см. рис. XI-6).
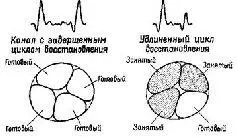
Рис. XI-6. Объяснение значения удлинения цикла восстановления возбудимости в афферентном канале после электрической стимуляции мозга. Эксперимент состоял в предъявлении пар стимулов, разделенных интервалами различной длительности. На график наносились изменения амплитуды ВП на второй стимул относительно амплитуды ВП на первый (в %) в зависимости от величины интервала, разделяющего эти стимулы (см. рис. XI-7). Предполагается, что, когда амплитуда ответа на второй стимул становится меньше амплитуды ответа на первый стимул, это означает, что афферентный канал в основном еще занят обработкой последствий действия первого стимула (показано на правой части рисунка). Обратите внимание на сходство этого рис. с рис. XI-5.
Читать дальшеИнтервал:
Закладка:








