К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Таким образом, во многих отношениях проблема поведения является обратной стороной проблемы Образа. Образ – это отражение внутри организма его окружения, а поведенческий акт – это отражение в окружающей среде того, что имеется внутри организма. Поведенческие акты – это достижение результатов; так, например, построение гнезда или написание книги совершается во внешнем мире, как оно было запланировано организмом, то есть соответствует его намерению или установке. То, как организуется поведенческий акт, и составляет предмет этого раздела нашей книги.
Вернемся еще раз к модели продуцирования слов, о которой мы говорили в начале этой книги. Предложение «Я вас люблю», произнесенное машиной, есть некий акт. Правда, здесь смоделирован только периферический голосовой акт, иными словами, электронная машина заменила собой уникальный голосовой аппарат человека. У человека настройка голосового инструмента должна осуществляться с помощью мозга. Тем не менее этот инструмент действует таким образом, что генерация даже такой сложной активности, как произнесение предложения, может осуществляться лишь посредством модификации двух мозговых процессов. Каким же образом тогда осуществляются эти модификации?
Я считаю, что организация действия есть в значительной мере управление рецепторными механизмами, содержащимися в сокращающейся ткани (в мышечных веретенах) или в сухожилиях, которые прикрепляют мышцы к костям и сочленениям. Эти рецепторы реагируют не только на сокращения мышц независимо от того, вызываются ли они внешними силами или нервными импульсами, возникающими в мозгу (через систему эфферентных волокон большого диаметра, известных под названием альфа-системы), но также и на возбуждения, приходящие к ним непосредственно через гамма-эфферентную систему волокон. Следовательно, непосредственная организация движения прямо связана с процессами, в которые вовлечены рецепторы, афферентные пути, идущие от этих рецепторов к спинному мозгу, и эффекторные пути, идущие от спинного мозга к сокращающимся мышцам и рецепторам (см. рис. V-2, V-6).
Всякий раз, когда этот процесс завершается стабильным ответом на стимуляцию, мы отождествляем его с рефлексом. В этой главе я впервые остановлюсь на некоторых интересных проблемах, касающихся осуществления рефлекса, который возникает благодаря наличию центрального контроля над рецепторами. Затем я перейду к детальному анализу мозговых механизмов управления функциями мышечных рецепторов.
В связи с существованием непосредственного контроля со стороны центральной нервной системы над рецепторными процессами посредством гамма-системы афферентных волокон, нейронную организацию рефлекса нельзя больше рассматривать как дугу или простую последовательность «стимул – реакция». Существует множество данных, свидетельствующих о том, что и сами рецепторы контролируются центральной нервной системой – особенно те из них, которые воспринимают мышечные сокращения. Этот центральный контролирующий механизм действует как система обратных связей и процессов предвосхищения, что, по существу, и определяет корригирующие возможности сервомеханизма. Получено так много доказательств справедливости этого положения, что Р. Гранит, один из первых исследователей как моторных, так и сенсорных механизмов, вынужден был недавно заявить следующее:
«… Что касается мотонейронов, то основные проблемы связаны не с рефлексами, хотя рефлексы всегда были и все еще остаются полезным средством анализа, необходимым в данной области знания. Основные проблемы касаются настройки или установки различных механизмов, посредством которых управляются мотонейроны – рефлекторно или как-либо иначе. Существует настройка мотонейронов с помощью нейромышечной интрафузальной системы гамма-волокон [находящейся в мышечных веретенах], а также посредством особых систем нейронных связей и нейрогормонов, действующих на альфа- и гамма-мотонейроны из высших уровней нервной системы» (Granit and Kellerth, 1967).
Относительно сервоконтроля движений многое стало известно после эксперимента, проведенного в духе нейропсихологических исследований и давшего парадоксальные результаты.
«Если мышца длительное время находится в состоянии сокращения под влиянием разрядов моторных импульсов, идущих от спинного мозга, удар тока, нанесенный на моторный нерв, приведет к возникновению судорожных всплесков на тензозаписи. Во время судорожного сокращения наблюдается прекращение разрядов моторных импульсов, идущих к мышце (рис. ХII-3). Это – период молчания. Это – рефлекторный акт (обсуждение этого вопроса см. Merton, 1951), который возникает, по-видимому, вследствие прекращения разрядов от мышечных веретен во время судороги. Именно эти разряды в обычных условиях вызывают рефлекс растяжения мышцы, и при их исчезновении мотонейроны замолкают. С точки зрения сервотеории, период молчания можно интерпретировать следующим образом: электрический ток размыкает замкнутую сервосистему и тем самым возбуждает поток добавочных моторных импульсов, который и вызывает сокращение мышцы; следовательно, механизм отрицательной обратной связи отключает спинальные моторные разряды до тех пор, пока мышца посредством расслабления не восстановит своей первоначальной длины.
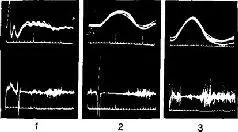
Рис. ХII-3. Периоды молчания в работе подошвенной мышцы человека, регистрация с помощью игольчатых электродов (нижняя запись). Под пяткой испытуемого закрепляется петля, и натяжение разгибателей щиколотки (верхняя запись) регистрируется при' давлении вниз на палец ноги. Время: 10 и 100 мсек; 5 суперпозиций в каждой записи; 1 – период молчания во время подергивания сухожилия, возникающего при ударе молоточком по ахиллесову сухожилию. При контакте молоточка с кожей возникает «всплеск» на кривой; 2 – запись во время рефлекса сокращения, вызванного электрическим раздражением афферентных волокон в подколенной ямке, 3 – запись во время судороги в латеральной части икроножной мышцы, вызванной стимуляцией выше этой мышцы. Обратите внимание, что подошвенная мышца сама по себе не возбуждается стимуляцией. Отметка времени (нижняя кривая на каждой записи) указывает 10 и 100 мсек (Granit, 1955).
Надо отметить, что отрицательная обратная связь не только нейтрализует эффекты изменений нагрузки или интенсивности добавочного потока импульсов, но делает работу мышц независимой от утомления или изменения синаптической возбудимости, точно так же, как усилитель, сконструированный по принципу обратной связи, не чувствителен к изменениям интенсивности поступающего тока или к напряжению электронных ламп. Стабильность – вот единственное требование, предъявляемое к системе обратных связей: в усилителе это сеть обратных связей со стабильным сопротивлением, в мышцах же это неутомляемые сенсорные органы. Как известно из работ Мэтгьюза, мышечные веретена как раз и обладают этими свойствами…
Читать дальшеИнтервал:
Закладка:








