К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Эта гипотеза в известной мере подтверждается экспериментами, недавно проведенными на нервно-мышечных приборах, которыми снабжена грудпая мышца. Хант (1951), в частности, наблюдал во время рефлекторного сокращения этой мышцы разряды моторных импульсов, посылаемых к интрафузальным мышцам (рецепторам веретен), предвосхищающие появление активности в основных мотонейронах» (Merton, 1953, р. 251-253).
Таким образом, произвольные движения, как правило, начинаются с активации интрафу зальных мышечных веретен.
Исключением из этого правила является тот случай, когда движения возникают неожиданно. Вследствие сравнительно малого диаметра гамма-волокон и большой длины всего пути скорость проведения слишком мала, чтобы объяснить внезапные мышечные сокращения. Однако подобные движения относительно просты и гамма-система быстро включается в контроль над этими движениями.
Следовательно, нельзя полностью игнорировать прямой путь к основным мотонейронам. Ограниченный объем предварительной информации, которая программирует процесс «сокращения – расслабления», может быть передан по этим путям, если интервал между началом и окончанием процесса будет достаточен для того, чтобы могла включиться в действие гамма-сервосистема. Работы Дж. Бермана и его сотрудников (Taub, Bacon and Berman, 1965) установили как верхнюю, так и нижнюю границы такого рода предпрограммирования движений. Берман использовал в эксперименте обезьяну, у которой спинной мозг ла всем его протяжении был полностью деаффектирован посредством удаления дорсальных корешков. Эта операция вызывает нарушение замкнутого сервомеханизма, обслуживающего движения. Однако после деафферентации у этих обезьян все же сохранялись условные реакции (сгибания конечностей). У человека же подобная деафферентация, широко практикуемая в настоящее время, чтобы снять боли и другие сенсорные нарушения, приводит к серьезным расстройствам автоматизированных движений, выполняемых соответствующей конечностью.
Итак, управление рефлекторными и интегрированными произвольными движениями независимо от того, вызываются ли они внешними или внутренними факторами, осуществляется в основном посредством настройки рецепторов мышечных веретен, включенных в гамма-сервосистему. Центральный контроль над движениями сводится, следовательно, к центральному контролю за рецепторными процессами. Эту проблему мы уже обсуждали во второй части, более подробно рассмотрим ее в четвертой части книги. Теперь мы сформулируем парадокс, который не укладывается в рамки узкого бихевиоризма: даже относительно простые, доступные для наблюдения двигательные механизмы связаны с рецепторными процессами, регулируемыми центральной нервной системой.
Признав факт существования произвольных движений, мы должны ответить на вопрос: как же они осуществляются? Ответ на этот вопрос можно дать на основе анализа процессов предвосхищения, с одной стороны, и раскрытия сущности предпрограммирующих процессов, подготавливающих результаты ряда движений прежде, чем они будут реализованы, – с другой.
Вспомним, что в гл. V указывалось, что для процессов предвосхищения необходим только приблизительный план, черновой набросок тех операций, которые должны быть выполнены. Как только будет выработан такой приблизительный план движений, дальнейшую их отшлифовку может взять на себя обычная сервосистема обратных связей. На основании нейро-анатомических и нейрофизиологических данных можно допустить, что некоторые аппараты нервной системы идеально приспособлены для обеспечения этой общей планирующей функции; и, как показывают результаты поведенческих экспериментов, эти функции действительно осуществляются такими аппаратами. Центральным звеном этой управляющей системы являются филогенетически новейшая часть мозжечка, эта удивительная структура, панорама которой открывается взору лишь при определенном сечении (рис. XII-9, XII-10).
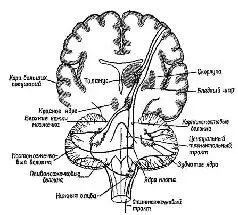
Рис. XII-9. Срединное сеченне через полушария и ствол мозга, на котором видно располошение и главные связи мозжечка Модифицированная схема Крига (1966).
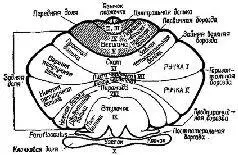
Рис. XII-10. Схематическое изображение борозд и долек мозжечка (Lar-sell, 1951; Jansen, Brodal, 1958; Angevine et al., 1961). Части мозжечка, расположенные сзади по отношению к постлатеральной борозде, составляют клочок (archicerebellum), а части мозжечка, расположенные спереди от первичной борозды, составляют переднюю дольку (paleocerebellum). Neoce-rebellum расположен между первичной и постлатеральной бороздами. Римские цифры обозначают только разделы червя мозжечка (Truex, Carpenter, 1969).
Как уже отмечалось выше, передняя часть мозжечка развилась при переходе морских позвоночных к наземному образу жизни. До этого момента мозжечок функционировал главным образом как дополнительный вестибулярный контролирующий механизм. Рыбы поддерживают ориентацию в соответствии с осью своего тела; их движения начинаются от этой основной линии. Амфибия и другие земноводные имеют дополнительную основную линию, это земля под ними, «низ», определяемый посредством гравитационных сил. Поэтому для организма становятся полезными новые вертикальные ориентиры позы и вследствие этого развивается передняя часть мозжечка. Наконец, приматы в известной мере освобождаются от гравитационной основной линии благодаря развитию двигательных навыков и приспособляются к древесному образу жизни и прямой позе. Теперь полушария мозжечка становятся доминирующими. У человека повреждение этих отделов мозга ведет к нарушению координации движений, к грубым промахам при слежении за целью и к атаксии при выполнении произвольных движений.
К счастью для нейрофизиологии, полушария мозжечка имеют удивительно гомогенное строение (рис. XII-11) и поэтому относительно просты для анализа. Их основными связями являются: 1) связи через спинной мозг со структурами, обеспечивающими афферентацию движений; 2) связи с корой больших полушарий (рис. ХП-9). Благодаря этим связям полушария мозжечка могут функционировать, как компаратор, сопоставляющий сигналы, поступающие от коры больших полушарий, с сигналами, идущими от периферии.
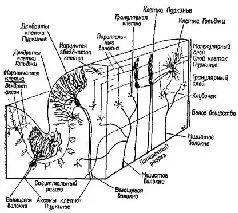
Рис. XII-11. Схематическое изображение коры мозжечка в сагиттальной и поперечной плоскостях, показывающее расположение клеток и волокон. Ср. с рис. XII-12, где дано изображение тех же клеток и волокон в поперечном разрезе (Truex, Carpenter, 1969).
Читать дальшеИнтервал:
Закладка:








