К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Статическое состояние функций мышцы называется ее тонусом. Однако тонус – это не величина спастичности или вялости сократительной ткани. Это, скорее, состояние готовности всего нейромышечного аппарата к действию, предварительное условие всякого действия.
Центральными структурами с замкнутыми системами связей, участвующими в регуляции тонической мышечной активности, являются базальные ганглии (рис. XII-6) и передняя часть мозжечка (рис. ХII-10). Поражение этих структур заметно изменяет состояние готовности мышечного аппарата: так, удаление переднего отдела мозжечка ведет к вялости, потере мышечного тонуса, а удаление базальных ганглиев – к полной обездвиженности, невыразительности мимики, маске Паркинсона и тремору конечностей, когда последние находятся в состоянии покоя. Филогенетически эти структуры мозга развивались, когда позвоночные впервые приспособились к передвижению по земле. Таким образом, эти структуры интимно участвуют в механизмах, регуляции позы, создающих то состояние готовности к ответу, тот фон, на котором может начаться дискретное действие.
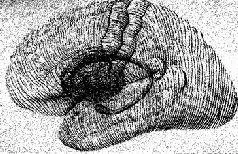
Рис. ХII-6. Расположение базальных ганглиев внутри больших полушарий головного мозга (Krieg, 1966).
Регуляция мышечной активности с помощью замкнутых кольцевых связей базальных ганглиев и передних отделов мозжечка была предметом большого числа исследований, однако в этом вопросе до сих пор нет полной ясности. По-видимому, в этих процессах регуляции участвует прежде всего гамма-система, хотя, как уже говорилось выше, альфа- и гамма-влияния участвуют также в интеграции движений. Наиболее существенные характеристики системы в значительной мере зависят от количественного и временного соотношения тех реципрокных взаимосвязей, которые установились между альфа- и гамма-активностью, а также от пространственной координации повторяющихся наборов сигналов в данной системе (рис. XII-7 и ХII-8). Эти характеристики трудно определить в экспериментальных ситуациях, в которых обычно применяются анестезирующие средства, что делает бесполезными попытки воспроизвести естественные механизмы регуляции позы. Тем не менее современные знания убеждают в том, что модуляция состояний готовности в мышечном аппарате осуществляется сервосистемой «базальные ганглии – передние отделы мозжечка» прежде всего посредством угашения непрерывных колебаний, присущих периферическому сервомеханизму с отрицательными обратными связями – механизму, определяющему рефлекторный акт. Это угашс-ние колебаний, вероятно, возникает благодаря пространственной координации, торможению и совместному действию реципрокных аппаратов различных сервомеханизмов. С помощью этих механизмов тоническое фоновое состояние моторной системы приобретает изменчивость и гибкость.
Рис. XII-7. Торможение разряда мышечного веретена при раздражении передней доли мозжечка. Верхние кривые на каждой записи показывают разряд веретена, распространяющийся вверх от черты, и артефакты от применения стимула, которые направлены вниз. Нижние кривые в каждой записи – миограммы. Децеребрированное животное. Икроножная мышца. Начальное давление – 66 гр. Миограмма записана при максимальном усилении (см. запись 12), за исключением 1-й записи, в которой сокращение (клони-ческое) в ответ на единичное раздражение электрическим током икроножного нерва выявляет молчащий период; 2-4 – контрольные записи перед стимуляцией; 5-10 – во время стимуляции мозжечка током 140 имп/сек при длительности импульса – 1 мсек, в течение 26 мсек; 5-7 – через 18-20 сек; 8-10 – непосредственно после прекращения стимуляции. Обратите внимание на падение частоты разрядов веретена от 20 имп/сек до нерегулярных разрядов порядка 5 имп/сек (Granit, Kaada, 1952; Granit, 1955),
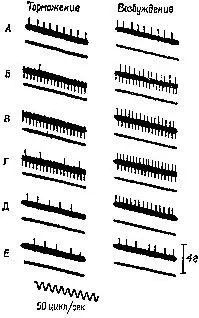
Рис. XII-8. Записи активности веретена подошвенной мышцы. Влияние стимуляции тормозных и активирующих участков ба-зальных ганглиев и других структур ствола мозга. Как и на рис. ХП-7, верхняя линия иа каждой записи показывает разряд веретена (подъем линий вверх) и артефакт от стимуляции (опускание линий вниз). А – средняя линия; В – первая запись во время стимуляции (обратите внимание на артефакты при электрическом раздражении) ; В – во время стимуляции; Г – последняя запись перед прекращением стимуляции; Д, Е – сразу же после прекращения стимуляции. На нижней кривой – миограмма. Начальное давление – 55 гр. Кошка под дозированным хлоролозовым наркозом (Eldred, Granit, Merton, 1953; Granit, 1955),
Сервоконтроль имеет и другой аспект. Иногда сервомеханизм не только регулирует движение «на выходе», но и создает предварительную установку к следующему этапу, то есть обеспечивает то предшествующее двигательной активности состояние, которое нуждается в стабилизирующих влияниях сервомеханизма. Эти процессы предвосхищения (рис. XII-5) широко распространены в моторной системе и обозначаются как произвольные, волевые или преднамеренные движения и действия. Современный уровень разработки проблем, связанных с произвольной активностью, еще не дает возможности понять механизмы нервного контроля поведения; хорошим началом в этом отношении могут служить исследования гамма-системы. Приведем еще одно-высказывание Мертона.
«Что же происходит во время произвольного сокращения? Легко осуществить произвольное сокращение мышц при устойчивом уровне двигательной активности; эти движения совершаются даже быстрее, чем самые быстрые судорожные сокращения. Скорость возникновения небольшого судорожного сокращения мышц лишь в малой степени соответствует скорости произвольного усиления. Здесь возникает очень интересная проблема: если во время такого усиленного сокращения мышц сервомеханизм будет продолжать действовать так же, как прежде, то он явно будет способствовать противоположному эффекту. Сразу же после начала произвольного сокращения мышечное веретено будет укорачиваться и возникающее в результате этого прекращение возбуждающих афферентных импульсов будет интенсивно подавлять мотонейроны… как если бы сокращение мышцы вызывалось небольшой стимуляцией двигательного нерва. Это значит, что дополнительные моторные разряды, возникающие при возбуждении мотонейронов спинного мозга, должны вызывать гот же самый эффект в замкнутой сервосистеме, что и импульсы, возникающие вокруг этой системы в их аксонах под влиянием электрического тока. В любом случае сервомеханизм должен энергично действовать, чтобы прекратить эти механические эффекты. Если эти выводы из экспериментов верны, то это означает, что во время произвольного сокращения следует преодолеть или устранить сопротивление, оказываемое сервосистемой… необходимость возбуждения основных мотонейронов, связанных с интрафузальными мышечными веретенами посредством нисходчщих импульсов, идущих из спинного мозга, исчезает. Возбуждение целиком поступает к небольшим мотонейронам (рецепторам мышечных веретен), н сервомеханизм запускает затем в действие основные мышцы. Таким образом, в произвольном сокращении мышцы участвует тот самый механизм, который обеспечивает постоянное фоновое сокращение, а именно: сервомеханизм, поддерживающий в мышце такой же ритм, как и в ее веретенах; следовательно, мышца находится под точно таким же стабилизирующим влиянием обратных связей, как и мышечные веретена. Предположение, что возбуждение поступает по основным мотонейропам, явно ошибочно, поскольку именно этот тип вмешательства, замкнутая система обратных связей и должна нейтрализовать. Замкнутая система наилучшим образом активируется в результате изменения режима работы ее «гасящих» детекторов (null-detektor).
Читать дальшеИнтервал:
Закладка:








