К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Как мы уже говорили, наблюдения Бернштейна показывают, что единство действия может быть достигнуто независимо от частных видов движения или количества сокращений каких-либо частных мышц или групп мышц. Действие может осуществляться лишь потому, что движения согласуются с «полем внешних сил», вовлеченных в действие. Как мы уже отмечали при рассмотрении гамма-эфферентной сервосистемы, существенные изменения в этой системе компенсируются указанным механизмом, так что постоянно сохраняется состояние готовности к движению. Таким образом, стабильность состояния достигается за счет специальных приспособлений и компенсаций внешних сил, а представительство в коре больших полушарий этого механизма включает эти параметры. Фактически центральный нервный механизм и состоит в основном, из этих приспособлений и компенсаций; следовательно, центральное представительство должно быть организовано таким образом, чтобы быть «зеркальным отражением» поля внешних сил (аналогичный вывод был сделан и в гл. XII).
Явное доказательство того, что нейроны моторной коры – особенно те, от которых идут пути на периферию, – действительно чувствительны к воздействиям, вызывающим мышечные сокращения, было получено в серии других остроумных опытов. В этих экспериментах Э. Эвартс (1967, рис. XIII-6) обучал обезьян работать рычагами, к которым были подвешены различные грузы, чтобы затруднить движения. Когда обезьяны научались двигать рычаг, в момент выполнения задания регистрировалась активность нейронов, расположенных в моторной коре. Опыты показали, что эти нейроны активируются до внешних признаков начала движения (по миографической записи). Тщательный анализ установил, что эта электрическая активность зависит от величины необходимых усилий, а не от величины смещения рычага. Более того, нейроны реагировали главным образом на изменение требуемой силы, хотя не исключено, что некоторая активность связана и с величиной необходимого усилия (рис. XIII-7).
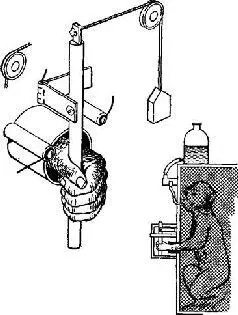
Рис. XIII-6. Аппарат для изучения поведения обезьяны с одновременной регистрацией моторного эффекта в клетках прецент-ральной коры. Левая рука обезьяны просовывается через отверстие в прозрачной перегородке, прикрепленной к передней стенке клетки. Чтобы получить фруктовый сок в качестве вознаграждения, обезьяна должна схватить вертикальный стержень, присоединенный к шарниру, и передвигать его взад и вперед между двумя упорами. Чтобы сработал соленоидный приемник, который «выдает» фруктовый сок, интервал между соприкосновениями стержня с контактами должен составлять 400-700 мсек и предварительное движение в другом направлении также должно совершаться в это же время (Evarts, 1968; см. Милнер, 1973).
Тот факт, что моторная кора предвосхищает параметры усилия, очень важен. Поскольку рефлексы организуются сервомеханизмами, центральное представительство этих рефлексов составляет не запись длины или напряжения мышц, а запись параметров приспособлений и компенсаций, соответствующих тем изменениям внешних сил, которые вовлечены в движение. Вследствие конвергенции этих процессов в центральном представительстве движений формируется образ не только прежних и текущих изменений внешних сил, воздействующих на систему, но – благодаря быстродействующим расчетам движений, совершающимся в мозжечке, – и тех изменений, которые должны произойти при продолжении двигательной активности.

Рис. XIII-7. Активность клетки моторной коры обезьяны в условиях, изображенных на рис. XIII-6. Показана активность клетки при трех видах нагрузки. Использовались следующие три вида нагрузки: 400 г на сгибатели (верхняя запись), 400 г на разгибатели (нижняя запись) и без нагрузки (средняя запись). Видно, что активность нейрона значительно возрастает при нагрузке на сгибатели, то есть при увеличении силы сгибания. Напротив, клетка почти полностью «замолкает» в те периоды, когда движение совершается при нагрузке на разгибатели (Evarts, 1967).
Таким образом, моторную кору можно рассматривать как сенсорную по отношению к действию. Она участвует в пространственной модуляции состояний готовности благодаря своим связям с базальными ганглиями и с механизмом быстродействующего-расчета параметров конечного эффекта действия, осуществляющегося при участии замкнутой мозжечковой системы. Формирование Образа результата зависит – как и всюду в коре – от того, что мы называем «нейронным топографическим представительством», то есть «от в высшей степени избирательного протекания активности в горизонтально расположенных корковых сетях, которые возбуждают и тормозят кортикофугальные нейроны» (Phillips, 1965). То, что это моторное представительство действительно во многих отношениях сходно с тем, которое формируется в сенсорных зонах мозговой коры, может быть установлено при исследовании рецептивных полей нейронов. Результаты этих экспериментов (Welt et al., 1967, табл. XIII-1) показали, что «сенсорная конвергенция в моторной (или сенсорной) коре накладывается на топографически сходные выходные структуры в виде радиальных рядов, диаметр которых, как оказалось, равен 0,1-0,4 мм. Таким образом, нейроны с фиксированными локальными рецептивными полями образуют радиально ориентированную структуру (референтную систему) по отношению к одним и тем же периферическим выходам… Вперемежку с этими клетками, имеющими локальные поля и составляющими 3/4 общей массы клеток, расположены нейроны с широкими, вертикально ориентированными или лабильными полями, которые перекрывают локальные поля» (стр. 285). Здесь, в моторной коре обнаружены явления, которые еще надо найти в зрительной коре (Теперь эти факты уже получены (Spinelli, Pribram and Bridgman, 1971).).
Таблица XIII-1

Сравнивались в простом рандомизированном плане антидромные латентные периоды реакции 88 корковых клеток, сгруппированных по типам рецептивных полей -Ответы группы «лабильных» клеток отличались от ответов обеих групп ‹Фикси1 рованных» клеток более чем на 1% not-критерию в предположении равенства групповых дисперсии.
s. d.- стандартное отклонение
S. Е.- средняя ошибка
См. описание сходства роли лабильных клеток и чувствительных клеток узко-яаправленного действия в корковых структурах Вернера (рис, VH-7), основанное/на -концепции Т-О-Т-Е (Welt et. al. 1967).
Читать дальшеИнтервал:
Закладка:








