К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Эту дилемму разрешил наш сотрудник Л. Гербрандт посредством простого наблюдения. Он показал, что амплитуда ответов на электрическую стимуляцию зрительной системы зависит от уровня внимания обезьяны во время опыта. Когда обезьяну сажали в закрытый ящик, реакция была слабой. Когда же ящик открывали и обезьяна получала возможность оглядываться по сторонам, реакция усиливалась. Более того, раздражение нижне-височной коры могло превращать слабый ответ, полученный в закрытом ящике, в сильный, но оно не оказывало влияния на «ильный ответ. И наконец, используя величину зтого вызванного при помощи электрической стимуляции ответа в качестве контрольного, Гербрандт мог предсказать, повлияет ли раздражение лижневисочной коры в условиях закрытого ящика на функцию восстановления возбудимости зрительной системы или нет.
В случаях, когда обезьяна была внимательна, ранее полученные нами .результаты не наблюдались. Когда же обезьяна начинала «скучать», клевать носом и т. д., влияние электрического раздражения на функцию восстановления было очевидным. В наших первоначальных экспериментах мы по заведенному порядку проделывали длительные ежедневные процедуры: парные вспышки, парные щелчки, комбинации щелчок – вспышка и вспышка – щелчок, вспышки по определенному шаблону и т. д., причем все это предъявлялось в определенном порядке, изо дня в день, из недели в неделю. Не только обезьяны, но и мы со Спинелли, проводившие эти опыты, разочаровались в этом рутинном порядке опытов. Спинелли и я поочередно следили за двумя своими обезьянами, чтобы они не заснули. Мы наблюдали за обезьяной через смотровое отверстие и, если она начинала дремать, тихонько постукивали по стенке. Если начинал дремать исследователь, мы с той же целью слегка сдвигали табурет, на котором он сидел Короче говоря, наши результаты объяснялись тем, что обезьяны не были внимательны. Впоследствии группы исследователей, изучавшие только цикл восстановления возбудимости у обезьян, и работавшие с обезьянами в новой для них ситуации, Kirja их явно интересовала процедура опыта, получали иные результаты. Только тогда, когда из-за повторения ситуация наскучивала и нам и обезьяне, вновь появлялись интересовавшие нас эффекты (Gerbrandt, Spinelli and Pribram, 1970, рис. XVII-9).
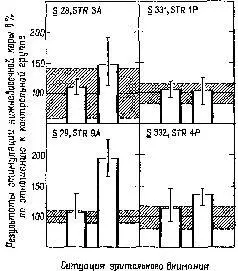
Рис. XVII-9. Влияние стимуляции нижневисочной коры на величину ответов, возникающих в зрительной коре при раздражении латерального коленчатого тела. Отметьте, что только в ситуации закрытого ящика (правая часть рис.) у трех обезьян из четырех имеется эффект, который превосходит дисперсию (затемненные участки), наблюдающуюся при отсутствии стимуляции (Gerbrandt, Spinelli and Pribram, 1970).
Эти эксперименты, вместе взятые, показывают, что результаты электрической стимуляции нижневисочной коры и результаты, получаемые, когда обезьяна «внимательна», сходны и что эти два процесса характеризуются высоким уровнем конвергенции на каком-то конечном механизме. Поэтому логично предположить, что процесс внимания предполагает влияние, оказываемое нижневисочной ассоциативной корой на механизм входа. Посредством этого влияния внимание может изменять временные параметры тормозных взаимодействий в фильтрах, а следовательно, и характеристику Образа, вызванного любым данным сигналом.
Таким образом, влияние височной доли на систему входа представляет собой частный случай и результат какого-то более общего процесса, посредством которого происходит усиление-определенных операций. Каким же образом тогда этот механизм: влияет на дифференцировку, на процесс узнавания?
Многочисленные исследования показали, что узнавание предполагает идентификацию признаков, различающих объекты. На этом основывается имитация этого процесса на ЭВМ при составлении программ. Открытие нейронов, чувствительных к определенным признакам, то есть способных избирательно реагировать на линии, наклонные под разным углом, движение, цвет и т. д., стало поэтому считаться решением проблемы узнавания. Не умаляя значения этого открытия, мы показали в предыдущих главах, что нельзя объяснить все проблемы восприятия наличием механизма детекции признаков. Позвольте мне развить эту точку зрения, высказав предположение, что проблема узнавания, завися-щая от памяти, также останется неразрешенной, если мы будем основываться только на детекции признаков. Поэтому я разграничу детекцию признаков и их опознание. Детекторы признаков являются необходимым звеном нервного аппарата и не могут быть радикально модифицированы в результате накопления опыта, если они должны выполнять задачу выделения признаков.
Поэтому детекторы признаков – это стабильные, «встроенные», врожденные элементы входных систем, которые проводят предварительную обработку сигналов, до того как они подвергнутся дальнейшей обработке. Напротив, механизм идентификации признаков, на котором основывается узнавание, должен по необходимости быть гибким и поддающимся модификациям в процессе приобретения опыта. Однако в силу непосредственности узнавания при предварительной обработке доходящей до мозга информации происходит одновременно как выделение признаков, так и процесс их опознания. Это участие двух разных процессов и привело к смешению этих двух механизмов и к предположению, что детекторы признаков целиком выполняют эту работу.
Детекторы признаков воспринимаемых объектов играют довольно-таки ограниченную роль. Полагают, что они наряду с другими механизмами обеспечивают необходимый уровень, фон, на котором возникают другие, более подвижные конфигурации нервных событий. Они представляют собой постоянные элементы, «основу», на которой возникает узор микроструктурных изменений, чувствительных к влиянию приобретаемого опыта. В любой данный момент эта «основа» подвергает обработке воздействующие на нее нервные сигналы, то есть подготавливает их к переключению на последующие клеточные элементы. Процесс обработки не оказывает влияния на саму «основу», но добавляет к ней еще один новый узор – еще одну нить.
Ряд таких механизмов, посредством которых опыт может оказывать влияние на нервную систему, был детально описан в гл. II и XIV. Эти механизмы, по-видимому, могут настраивать, изменять действие тех элементов, которые составляют экран. Небольшие изменения времени восстановления возбудимости нервных элементов, вызываемые, например, стимуляцией нижневисочной коры, могли бы, если бы они стали постоянными, изменять характеристики ответов элементов системы, пока они не будут «настроены» особым образом для восприятия только данной конфигурации пейронной микроструктуры. Спинелли ( 1970, см . рис. VII-8) имитировал в программе ЭВМ подобную систему, названную им «Оккам». Эта программа может «узнавать» форму волны, если она применялась несколько раз, и может опознавать данную структуру даже по ее части. Кроме того, эта программа может различать десятки форм волны.
Читать дальшеИнтервал:
Закладка:








