К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии

- Название:ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
К.ПРИБРАМ - ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии краткое содержание
Под редакцией и с предисловием
действительного члена АПН СССР
А. Р. Лурия
Издательство «Прогресс» Москва 1975
Редакция литературы по философии
Перевод на русский язык с изменениями. "Прогресс"
ЯЗЫКИ МОЗГА Экспериментальные парадоксы и принципы нейропсихологии - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Можно, пожалуй, считать, что нервный механизм, посредством которого осуществляется опознание, действует сходным образом. Обычно организованные элементы фильтра, колонки кортикальных клеток более или менее связаны между собой посредством элементов, чувствительных к направлению. Напомним, что в гл. VII высказывалось предположение, что чувствительность к направлению рецептивных полей корковых клеток является определяющим фактором для соседних клеток, превращая таким образом кортикальную колонку в структуру,, включающую определенные координаты, или реестр. Латеральное-торможение строит этот реестр путем разделения организованных в определенную систему элементов. Каждый модуль этой системы, каждый реестр можно представить себе как диполь,, поляризующийся при помощи входных сигналов. Многое говорит в пользу существования электрических диполей в коре (см. Elul, 1964; Barrett, 1969). Электрический диполь мог бы образоваться из тех изменений в молекулярной структуре, которые обсуждались в гл. II и VIII. Эти структурные изменения можно-измерить при помощи ядерной магнитно-резонансной спектроскопии и изучения дифракции рентгеновских лучей, поскольку каждая структура резонирует на своей частоте. Известно, что макромолекулы типа липопротеинов и глюкоидов, образующие-синаггхические и дендритные мембраны в микроструктурах мозга,, подвержены таким изменениям. Всякий раз, когда нервный сигнал проходит через такую мембрану, эти изменения имеют тенденцию-стабилизироваться, упорядочивая молекулярные структуры, пока длится сигнал и в течение некоторого ограниченного времени после него. Если эти временно стабилизирующиеся изменения достаточно" обширны, они будут вызывать электрическую поляризацию микроструктур. Эта поляризация возрастет, когда вследствие латерального торможения влияние каждого диполя на соседний становится минимальным. Без таких тормозных взаимодействий действие диполей имело бы тенденцию к ослаблению, подобно тому, как полюса магнитов, поставленные параллельно, имеют тенденцию нарушать упорядочивание. Поэтому в результате влияния раздражения височной коры на латеральное торможение действие-диполей при входном сигнале возрастает (см. рис. XVII-10), причем это возрастание обеспечивает постоянство связей между входным сигналом и организованными элементами системы. Эти связи составляют мелкие корковые функциональные единицы, обеспечивая тем самым более легкую адаптацию каждого элемента к своему входному сигналу. Как указывал Росс Эшби (1960, рис. XVI1-10), полностью взаимосвязанная система ультрастабильна, что затрудняет адаптацию. Модификация может происходить только путем «разделения системы на части» при постоянстве связей.
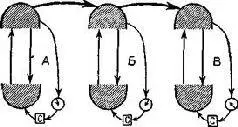
Рис. XVII-10. Схема установления устойчивого состояния в гомеостате Эшби. Верхние полукруги изображают вариации стимулов; нижние – вариации нервных ответов. С – выключатель селектора, стрелка-монитор. Обратите внимание на сходство этой схемы со схемой Т -О -Т -Е (Ashby, 1960).
Такая система диполей, структур нейронных реестров имеег определенные преимущества, потому что она представляет собой, параллельно обрабатывающий механизм, связанный с содержанием. Элементы такой системы действуют одновременно, а непоследовательно; место хранения информации теряет свое значение. Предварительная обработка и модификация, основанные на опыте, происходят бок о бок и практически одновременно. Короче говоря, некоторые части механизма предварительной обработки изменяются в процессе этой обработки. Нервный механизм самоорганизуется.
Подведем итог. В реальной жизни нелегко разграничить перцептивный процесс формирования Образа и познавательный процесс его опознания. Мы опознаем то, что ощущаем как значительное, почти в тот же момент, когда воспринимаем это посредством своих органов чувств. Но для целей научного анализа, особенно если нас интересует механизм, важно расчленить, как мы и сделали в этих главах, различные участвующие в этом процессы. Поскольку в итоге мы выясняем, что нервные механизмы этих процессов в действительности тесно взаимосвязаны, мы приходим к более полному пониманию того, каким образом возникает единство психологического процесса. Когда это единство ясно выражено, знак опознается или актуализируется, но эта актуализация (как и все действия) должна осуществляться последовательно. Любое мгновенное формирование знака поэтому не является полным – знак извлекает значение только из целостного Образа и только тогда может заменить его.
Нам осталось обсудить последний вопрос: как некоторые аспекты Образов обретают смысл, становятся значимыми? В экспериментах по выработке дифференцировок у обезьян какое-либо изображение получает смысл, становится знаком в результате поведения самой обезьяны. При выборе используется нервный механизм, необходимый для совершения действия. Тот факт, что проводящие пути от нижневисочной области коры, которая оказывает влияние на зрительное внимание, проходят через двигательные отделы мозга, обеспечивает структурную базу для взаимодеиствия двигательных и сенсорных процессов и для влияния-Образов результата на Образы восприятия. Поэтому значение данных сигналов создается посредством действия. Именно, этот активный процесс образования знака порождает значение: он является результатом перцептивного научения посредством подкрепления. Оказывается, что значимость сигналов зависит от механизма, который привлекает к ним внимание, от подкреплений и альтернатив (см. рис. XVII-11). Обезьяны, лишенные нижневисочной коры, выбирают из ограниченного числа альтернатив (при меньшей неопределенности); зрительный выбор является результатом четкого различения воздействующих агентов (Pribram, 19606, см . рис. XIX-11, XIX-12, Butter, 1968, рис. XVII-12). Если животное внимательно или если нижневисочная кора подвергается электрической стимуляции, происходит замедление восстановления сенсорных систем (рис. XI-5, XI-6). Такое замедление уменьшает избыточность поступающей информации. Таким образом, данный ряд сигналов начинает входить в более мелкие части нейронных голограмм. Когда для реконструкции Образов используются маленькие части физической голограммы, глубина поля возрастает и большая часть Образа попадает в фокус, хотя и ценой некоторых небольших потерь в деталях. Работа этого механизма весьма напоминает укорочение фокусного расстояния транслокаторного фотообъектива – при этом в фокусированное поле зрения попадает большая площадь. Были получены прямые доказательства наличия такого механизма у человека. Измерения рецептивных полей отдельных элементов в зрительной коре человека показывают, что константность восприятия величины достигается здесь при помощи процесса, поддерживающего постоянный угол относительно точки фиксации взора в геометрическом центре рецептивного поля (Marg and Adams, 1970). Так, когда я хочу рассмотреть большую часть какой-либо сцены, я укорачиваю фокусное расстояние. Парадоксально, что, когда затем меня начинает интересовать тот или иной аспект этой сцены, соответствующие признаки воспринимаемого выделяются и становятся свободными от влияния побочных деталей Когда же, напротив, я намереваюсь уточнить какой-то ограниченный сегмент данной сцены и исключить все остальное, я удлиняю фокусное расстояние. Сегмент, находящийся в фокусе, лишается, таким образом, контекста, с которым первоначально был связан, вследствие чего этот сегмент может получать разные значения в зависимости от цели или установки наблюдателя.
Читать дальшеИнтервал:
Закладка:








