Петр Анохин - Проблемы лечебного голодания. Клинико-экспериментальные исследования

- Название:Проблемы лечебного голодания. Клинико-экспериментальные исследования
- Автор:
- Жанр:
- Издательство:Типография №1 Росглавполиграфкомбината Комитета по печати при Совете Министров РСФСР
- Год:1969
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Петр Анохин - Проблемы лечебного голодания. Клинико-экспериментальные исследования краткое содержание
Представленные в этом сборнике работы дают подробное описание клиники лечебного голодания, нейрофизиологии голода и насыщения, патофизиологии и биохимии голодания. Авторами описано большое количество клинических примеров излечения больных с помощью лечебного голодания от целого ряда психических и соматических заболеваний, в том числе считающихся «неизлечимыми» и трудно поддающимися лечению.
Сборник будет интересен врачам различных специальностей, психиатрам и психотерапевтам, патофизиологам и специалистам других медицинских и биологических наук, студентам высших и средне-специальных медицинских учебных заведений, искателям здорового образа жизни и всем остальным, кто глубоко интересуется лечебным голоданием, включая тех, кто уже имеет личный опыт этой методики, поскольку представленная информация имеет огромную научную ценность для понимания сути процессов, происходящих в процессе лечебного голодания.
Этот сборник вышел в 1969 году, но, несмотря на это, до сих пор распространен миф, что голодание свыше нескольких дней наносит непоправимый вред организму. Поэтому одна из главных задач этого тома — продемонстрировать физиологичность голодания и безвредность больших сроков при этой методике.
Проблемы лечебного голодания. Клинико-экспериментальные исследования - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Дальнейшее подтверждение эта точка зрения нашла в исследованиях, показавших, что различные наркотические вещества также избирательно действуют на системы корково-подкорковых взаимоотношений, вовлекаемых в формирование реакции того или иного биологического качества.
Так было показано, что такое наркотическое вещество как нембутал почти полностью блокирует все формы восходящих активирующих влияний со стороны подкорковых образований на кору мозга. В то же время другое наркотическое вещество — уретан, блокируя активацию коры, связанную с состоянием бодрствования, допускал развитие корковой активации вслед за нанесением болевого раздражения (I).
Все эти наблюдения позволили взамен представления о неспецифическом механизме восходящих активирующих влияний ретикулярной формации на кору мезга сформулировать представление о том, что подкорковые образования оказывают на кору мозга множественные восходящие активирующие влияния, каждое из которых соответствует определенной реакции того или иного биологического качества (П. К. Анохин, 1962) (2).
Такое представление позволило с новых позиций приступить к изучению корково-подкорковых механизмов «основных влечений» организма, одним из которых является состояние голода. Мы предположили, что пищевое возбуждение у голодных животных строится на основе своих специфических механизмов активации коры головного мозга со стороны подкорковых и пищевых центров.
В связи с этим мы поставили перед собой следующие основные задачи:
1. Изучить, каким образом пищевое возбуждение у голодных животных проявляется в электрической активности коры головного мозга?
2. Какие подкорковые образования определяют пищевое возбуждение животных?
3. Каковы функциональные взаимоотношения подкорковых центров с корой головного мозга у голодных животных?
4. Каким образом подкорковые образования вовлекают в свою деятельность элементы коры головного мозга, т. е. каковы те нейрофизиологические механизмы, на основе которых строится пищедобывательное поведение животных и человека?
Опыты были проведены на 283 кошках под уретановым наркозом. Уретан вводили внутрибрюшинно в 7,5% или 15% растворе из расчета 1,5 гр. на кг веса животного.
В процессе наших экспериментов было установлено, что уретан, как наркотическое вещество, избирательно блокируя состояние бодрствования, в то же время оставлял относительно интактным нейрофизиологические механизмы пищевых возбуждений. Все это позволяло исследовать центральные механизмы «голодных» возбуждений в «чистом виде», не осложненные возбуждениями других биологических модальностей.
Под уретановым наркозом у животных регистрировалась ЭЭГ. Отведение ЭЭГ осуществлялось при помощи стальных игольчатых электродов, которые вкалывали непосредственно в кость черепа.
В большинстве опытов ЭЭГ отводили униполярным способом. Индифферентный электрод помещали по средней линии над лобной пазухой.
Для записи электрической активности подкорковых образований мозга так же как и для воздействия на них использовали изолированные за исключением кончика специальные погружные нихромовые электроды. Ориентацию погружения электродов в подкорковые образования проводили согласно координатам стереотаксического атласа мозга кошки, составленного Джаспером и Айжмон-Марсаном (20).
ЭЭГ записывали на чернилопишущем 10-канальном электроэнцефалографе фирмы «Альвар-Электроник».
В ряде опытов в коре мозга регистрировали вызванные потенциалы, возникающие в ответ на электрическое раздражение седалищного нерва и на раздражение подкорковых гипоталамических структур.
Регистрация вызванных ответов в коре мозга осуществлялась па 4-канальном катодно-лучевом - осциллографе «Биофаз» фирмы «Альвар-Электроник» с полосой пропускания 0,5 гц до 10 кгц.
Электрическое раздражение осуществляли стимулятором «физиовар» с трансформаторным выходом.
В специальной серии опытов производили коагуляцию или поляризацию различных подкорковых образований. Это достигалось действием на них анода постоянного тока. Локализацию кончиков электродов в мозге определяли проекционным методом (26). В ряде опытов препараты окрашивались по методу Ниселя.
Электрическая активность коры головного мозга у животных в состоянии голода и после приема пищи
Для того, чтобы характеризовать, каким образом возбуждения подкорковых отделов пищевого центра выходят на «кору мозга, в первой серии экспериментов нами изучался характер электрической активности коры мозга у животных в различные сроки голодания и после приема пищи.
Опыты-показали, что у подавляющего числа животных под уретановым наркозом после 1—2-суточного голодания в передних отделах коры головного мозга регистрировалась высокочастотная низкоамплитудная электрическая активность (амплитуда колебаний 6—10 мкв, частота 20—25 кол/сек), которая резко отличалась от высокоамллитудной (50—80 мкв), относительно медленной (6—8 кол/сек) активности теменно-затылочных областей.
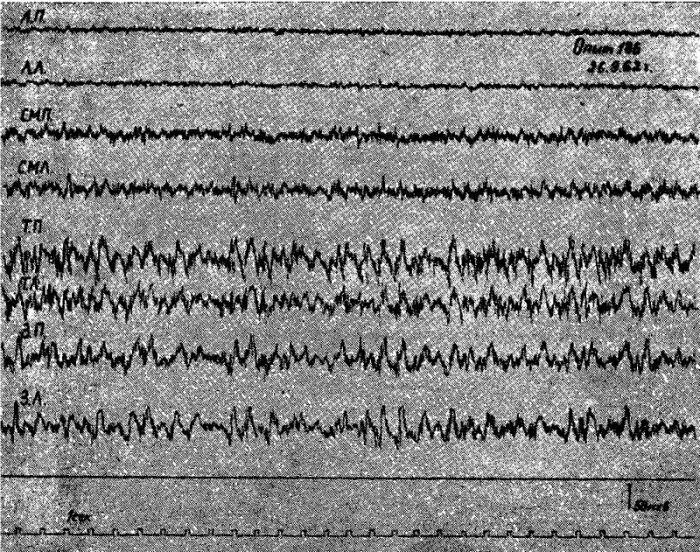
Рис. 1. Электрическая активность коры мозга кошки под уретановым наркозом после 2-суточного голодания.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая, СМП — сейсомоторная правая, CMЛ — сензомоторная левая, ТП — теменная правая, ТЛ — теменная левая, ЗП — затылочная правая, ЗЛ — затылочная левая обл. Отметка времени 1 сек.
Граница распространения но жоре головного мозга высокочастотной активности у животных после 1—й-суточного голодания была определена в специальных опытах. Такая активность в наиболее отчетливой форме наблюдалась над отделами мозга, расположенными впереди венечной борозды. Однако высокочастотная активность регистрировалась и в каудальных частях передних отделов мозга, в которых по мере продвижения в направлении теменных отделов постепенно нарастала высокоамплитудная активность. Задней границей распространения зоны высокочастотной низкоамплитудной ЭЭГ активности у животных после кратковременного голодания была область сильвиевых борозд.
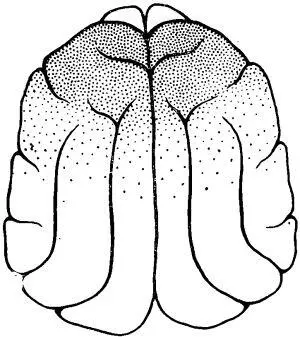
Рис. 2. Схема распространения под уретановым наркозом реакции, десинхронизации по коре мозга у кошки после 2-суточного голодания.
У накормленных перед опытом животных характер электрической активности коры мозга значительно отличался от активности голодных животных. У таких животных со всех отделах коры мозга регистрировалась медленная высокоамплитудная электрическая активность (амплитуда 50—80 мчсв, частота 3—5 кол/сек)
Читать дальшеИнтервал:
Закладка:








