Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
В-пятых, после работы рибозима-полимеразы образуется длинная двунитевая РНК, которая не может служить матрицей для следующего копирования. Чтобы продолжить копирование, надо как-то расплести ее на две отдельные нити. В клетках это делают специальные ферменты – хеликазы, расходующие энергию в виде АТФ на расплетание двойных спиралей, а специальные белки (они называются SSB – single-strand binding) связываются с одиночными нитями, чтобы они не слипались обратно. В раннем РНК-мире такой сложной системы, очевидно, быть не могло, и надо искать другие способы расплетания двунитевых РНК. Простейшим способом могут быть колебания температуры, которые используются в полимеразной цепной реакции (ПЦР) – лабораторном методе копирования ДНК в пробирке, при помощи одной полимеразы. В полимеразной цепной реакции построение вторых цепей ДНК идет при температуре 60–70 °C, а периодический короткий нагрев до 95 градусов разделяет двунитевые молекулы на отдельные нити. В условиях древней Земли подобные колебания температуры могут давать периодические выбросы кипятка из гейзеров или приливы на морском берегу. Однако при падении температуры две длинные нити РНК могут соединиться обратно. Для запуска нового цикла копирования нужно, чтобы концентрация затравок была в 100–1000 раз выше, чем длинных молекул. В ПЦР этого добиваются, добавляя большое количество искусственных затравок. В РНК-мире доступность затравок особенно критична при копировании РНК в липидных пузырьках, стенки которых пропускают только одиночные нуклеотиды, но и в растворе, и на минералах затравки все равно нужны.
Копирование РНК путем крупноблочной сборки
Стало понятно, что надо искать другие подходы к копированию РНК. Например, процесс копирования мог поначалу происходить путем сшивания концов коротких цепей, а не одиночных нуклеотидов. Обзор Карлоса Брионеса с соавторами (2009) суммирует главные открытия в этой области.
В этом сценарии первые РНК длиной 20–30 нуклеотидов образуются при случайной полимеризации на минералах. Многие из них имеют участки, комплементарные другой части молекулы, и образуют шпильки (рис. 9.3). Среди них были первые рибозимы, проводящие реакцию сшивания концов двух цепей РНК, под названием «лигазы». Они сшивали некоторые РНК друг с другом, что приводило к появлению, во-первых, более активных лигаз, а во-вторых, эти лигазы лучше узнавали те короткие РНК, из которых могли быть построены их собственные копии. По мере роста размеров и сложности РНК среди лигаз могли появиться и рибозимы-полимеразы.
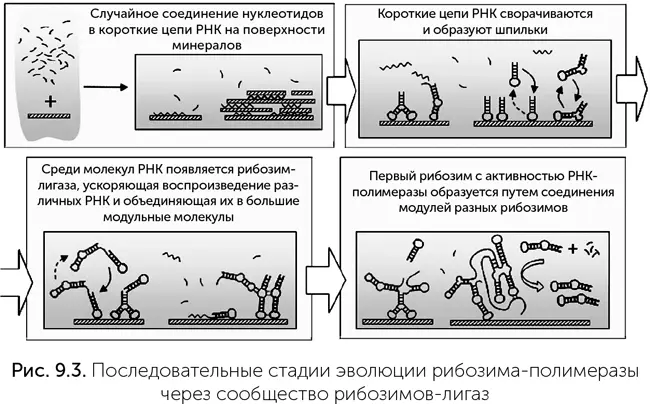
Такой сценарий решает часть проблем, которые были в сценарии случайного появления полимеразы. Во-первых, рибозимы-лигазы гораздо проще и короче, чем рибозимы-полимеразы: при комнатной температуре хорошо работают лигазы длиной 40–50 нуклеотидов, а во льду – даже более короткие фрагменты, вплоть до 29 нуклеотидов (Vlassov et al., 2004). Молекулы РНК таких размеров реально получить при случайной сшивке нуклеотидов без помощи рибозимов. Во-вторых, лигазы могут сшивать две разные функциональные молекулы РНК, образуя более крупные модульные рибозимы, – путь усложнения, малодоступный для рибозимов-полимераз. В-третьих, в экспериментах рибозимы-лигазы склонны образовывать сообщества, в которых разные лигазы ускоряют сборку друг друга, поддерживая более сложную и разнообразную систему, чем один рибозим-полимераза. Более того, такие сообщества выигрывают в конкуренции с эгоистичной лигазой, собирающей только свои копии. В экспериментах Трейси Линкольн и Джеральда Джойса пара лигаз собирала копии друг друга (Lincoln, Joyce, 2009) и удваивала свою численность менее чем за час, тогда как полимеразы не могут собрать свою копию и за сутки! Более того, добавив в систему несколько версий «сырья» – коротких цепочек РНК, они добились конкуренции разных вариантов рибозимов-лигаз и начала эволюции среди них. Единственный недостаток известных сообществ лигаз, не позволяющий признать их окончательным решением проблемы копирования в мире РНК, – это необходимые для них исходные материалы. Лигазы Линкольн и Джойса так же, как их аналоги, полученные другими учеными, требуют довольно длинных фрагментов РНК, около 20 нуклеотидов, для построения из них собственных копий. Если материал для лигаз поставляется случайным соединением нуклеотидов на глине, то вероятность получения нужного 20-нуклеотидного фрагмента РНК будет крайне мала – около одной триллионной. Лигазы, хорошо работающие с кусочками РНК в три – пять нуклеотидов, пока получить не удается.
Интересный вариант содружества лигаз был получен в работе Щепанского и Джойса (Sczepanski, Joyce, 2014). Они отступили от принципа хиральной чистоты и сделали рибозим из нуклеотидов с «левыми» молекулами рибозы, отбирая его на способность сшивать обычные РНК с «правой» рибозой. В этом случае звенья рибозима не образуют комплементарных пар с теми молекулами, которые он сшивает. Подобные взаимодействия приводят к застреванию молекул РНК в обычных рибозимах-полимеразах и лигазах и сильно замедляют их работу. «Левый» рибозим из 83 нуклеотидов оказался очень хорошей лигазой, способной построить свою зеркальную копию из 11 коротких фрагментов РНК по 7–11 нуклеотидов. Зеркальная копия, в свою очередь, так же эффективно строит исходный «левый» рибозим из «левых» коротких РНК. Такой способ копирования был назван «кросс-хиральная репликация». Как и некоторые другие лигазы, этот рибозим может по совместительству «подрабатывать» полимеразой: он достраивает РНК-затравку, присоединяя к ней до шести нуклеотидных звеньев, причем к «правой» затравке присоединяются только «правые» нуклеотиды из смеси, а к «левой» – «левые». Этот эксперимент заставляет задуматься об истории хиральной чистоты: может быть, мир РНК сначала использовал левые и правые нуклеотиды в равной мере, а хиральная чистота возникла позже, с переходом к белкам? Авторы надеются в ближайшие пару лет сделать из этой лигазы кросс-хиральную полимеразу, которая будет строить из отдельных нуклеотидов свои зеркальные отражения. Посмотрим, что у них получится.
Итак, ни рибозимы-полимеразы, ни рибозимы-лигазы, известные на сегодняшний день, не обеспечивают устойчивого копирования РНК из доступных в окружающей среде древней Земли компонентов – отдельных нуклеотидов и их цепочек длиной до пяти-восьми звеньев. Чтобы запустить копирование, а следовательно, конкуренцию и естественный отбор, необходимы какие-то другие способы копирования РНК.
РНК копируются внешними силами
Самопроизвольное соединение активированных нуклеотидов, с которого мы начали эту главу, может происходить не только на глине, но и на однонитевых молекулах РНК, работающих в качестве матрицы. Достижения и проблемы в этой области обобщены в обзоре Джека Шостака (Szostak, 2012).
Читать дальшеИнтервал:
Закладка:






