Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
В поре, открытой с обоих концов, поведение молекул несколько отличается. Если есть проток жидкости снизу вверх и нагрев с одной стороны, то в поре тоже происходят конвекция и термофорез. Молекулы РНК накапливаются на нижнем конце холодной стенки, но их поведение сильно зависит от длины. Нуклеотиды и короткие РНК слабее увлекаются термофорезом в нисходящий холодный поток и в итоге вымываются из поры с восходящим потоком, а РНК длиннее определенного порога (он зависит от скорости потока и размеров поры) накапливаются в ней. Это избирательное удержание длинных РНК может противостоять быстрому размножению коротких РНК и сохранять длинные молекулы в ряду поколений несмотря на то, что они копируются медленнее (Kreysing et al., 2015).
И в открытых, и в закрытых порах благодаря конвекции молекулы РНК последовательно попадают в горячие и холодные зоны. В горячей зоне двухцепочечная РНК может разделяться на одиночные цепи, которые смогут стать матрицами для следующего цикла копирования.
Мир шпилек
Изящное решение проблемы затравок было недавно предложено Александром Марковым ( http://macroevolution.livejournal.com/170098.html). Оно состоит в том, что в условиях неферментативного копирования РНК преимущества получают те последовательности, которые могут служить затравками для синтеза собственных копий (рис. 9.5). Такими свойствами обладают палиндромные последовательности РНК. Палиндромами называли слова или фразы, которые читаются одинаково в обе стороны («А роза упала на лапу Азора»). В случае РНК и ДНК палиндромной последовательностью называют такую, которая читается одинаково слева направо и – после замены нуклеотидов на комплементарные – справа налево. Например, последовательность GGACCUAGGUCC будет палиндромом.
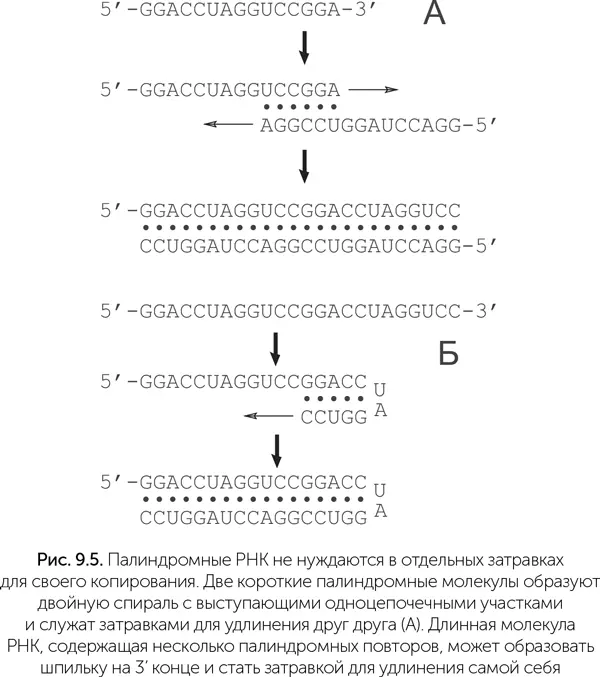
При случайном разрыве таких цепей РНК получаются короткие фрагменты, которые тоже могут служить затравками для удлинения своих родственников. В условиях неферментативного копирования РНК идея Маркова («Мир палиндромов») превращает необходимость затравок из проблемы в фактор естественного отбора, который начинается еще до появления первого активного рибозима, с молекулами РНК длиной 15–20 нуклеотидов. Причем низкая скорость неферментативного копирования здесь не страшна: достройка концов палиндромных РНК требует присоединения лишь 5–10 нуклеотидов за один цикл. Палиндромность первых РНК повышает вероятность появления рибозимов, так как палиндромные последовательности образуют множество шпилек (рис. 9.6).
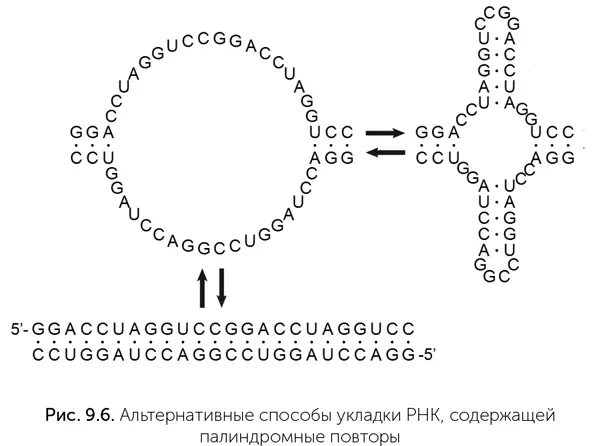
Следы такого способа роста структур РНК до сих пор видны в древнейшем реликте РНК-мира – молекулах транспортных РНК (тРНК). Эти молекулы длиной 76 нуклеотидов имеют форму клеверного листочка. «Стебелек» и каждый из «листиков» трилистника включают двуспиральные участки РНК. Три «листика» образуют шпильки, а стебелек заканчивается свободными концами цепи РНК. В последовательности тРНК давно были обнаружены внутренние повторы, допускающие другие варианты укладки молекулы, например, в виде одной длинной двухцепочечной шпильки с несколькими внутренними петлями. На основе этих повторов были предложены модели роста тРНК путем увеличения количества палиндромных сегментов молекулы (Rodin et al., 2011; DiGiulio, 2009).
Если у нас есть механизм абиогенного копирования РНК, хотя бы только коротких и палиндромных молекул, то среди них могут оказаться РНК с какой-то рибозимной активностью. Строго говоря, не только лигаза и полимераза способны ускорить накопление своих копий в такой системе. Например, рибозимы-экзонуклеазы, разрушающие молекулы РНК, могут быть полезными для их точного и быстрого копирования. Экзонуклеазы работают в клетках вместе с полимеразами и служат для отрезания неправильно присоединенных нуклеотидов, повышая точность копирования. Рибозим с такой активностью мог бы повышать точность неферментативного копирования РНК и косвенно – скорость копирования, потому что после неправильно присоединенного нуклеотида (не образующего комплементарную пару с матрицей) следующий нуклеотид присоединяется намного медленнее обычного. Кроме того, для увеличения количества собственных копий рибозим может вообще воздействовать не на копирование РНК, а на предшествующие шаги – превращение простых органических веществ в нуклеотиды. Иначе говоря, появляется обмен веществ – химические реакции малых молекул, происходящие под контролем ферментов. В следующей главе мы подробнее рассмотрим, на что способны рибозимы в процессе обмена веществ.
Глава 10
Витамины, аминокислоты и пептиды в РНК-мире
Рибозимы осваивают обмен веществ
Как только на Земле появляются молекулы РНК, которые как-то копируются, между ними начинается конкуренция. Одни молекулы РНК копируются быстрее, чем другие, и могут накапливаться в большем количестве. Любая новая особенность рибозима, повышающая вероятность его копирования, будет подхвачена естественным отбором.
В прошлой главе мы обсудили возможности и ограничения рибозимов-полимераз и лигаз, которые прямо участвуют в копировании РНК. Но это не единственные способы, которыми рибозим может влиять на количество своих копий и их эволюционную судьбу. Как справедливо замечено в обзоре Martin et al., 2015, путь от простой органики к РНК состоит из многих шагов, и ускорение любого из них при помощи рибозима будет выгодно для этого рибозима. Рибозимы в принципе могут как ускорять уже идущие реакции синтеза и активации нуклеотидов, например синтез Сазерленда, так и пускать в дело отходы и побочные продукты – свободные азотистые основания, рибозу и нуклеозиды, которые образуются при распаде РНК и нуклеотидов. Кроме того, если конкурирующие молекулы РНК находятся в лабиринте пор в минеральном осадке, то между соседями вероятно возникновение кооперации, когда каждый из них ускоряет копирование не только самого себя, но и молекул, находящихся рядом. В таком сообществе будет разделение труда, когда одни рибозимы копируют РНК, а другие занимаются производством нуклеотидов для них. Так появляется обмен веществ: химические реакции между малыми молекулами, проходящие под контролем соседних молекул; такие помощники-контроллеры получили название «ферменты».
Ресурсы, которые потребляет это сообщество РНК, можно рассортировать по сложности использования. Проще всего применять, конечно, готовые активированные нуклеотиды, такие как АТФ и ГТФ (гуанозинтрифосфат, у которого три фосфатные группы присоединены не к аденину, а к гуанину), – это одновременно и строительный материал для новой РНК, и источник энергии. Следующими после готовых активированных нуклеотидов будут нуклеотид-монофосфаты – готовые строительные блоки РНК, но без запаса энергии. Еще немного сложнее использовать нуклеозиды – чтобы пустить их в дело, надо добавить фосфатную группу. И нуклеотид-монофосфаты, и нуклеозиды неизбежно появлялись при распаде цепочек РНК.
Читать дальшеИнтервал:
Закладка:






