Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Многие вспомогательные компоненты репликативной вилки имеют общее происхождение у бактерий и архей: ДНК-лигазы, скользящий зажим, загрузчик скользящего зажима. РНКазы Н бактерий и архей тоже сходны и имеют общее происхождение, но не обязательно от LUCA. Этот же фермент есть и у ретровирусов. Он мог независимо попасть от них в клетки бактерий и архей.
Общее происхождение скользящего зажима и его погрузчика означает, что структура репликативной вилки, копирующей одновременно две цепи, могла существовать у LUCA – для более простых механизмов, например «катящегося кольца», скользящий зажим и ферменты его погрузки не требуются. Существование репликативной вилки указывает на большие геномные молекулы длиной более 100 000 пар нуклеотидов. Наличие у LUCA ДНК-лигазы и ДНК-зависимой РНК-полимеразы означает, что ДНК в какой-то форме у него уже была. Но принципиально разные ДНК-полимеразы и праймазы бактерий и архей объяснить труднее.
Возможны три основных объяснения:
1) LUCA имел обе версии системы репликации ДНК одновременно, бактерии унаследовали одну, археи – другую;
2) LUCA имел одну из современных систем репликации, в одной из двух линий потомков она была заменена на новую;
3) система репликации LUCA принципиально отличалась и от бактериальной, и от архейной, обе линии потомков ее заменили на современные варианты.
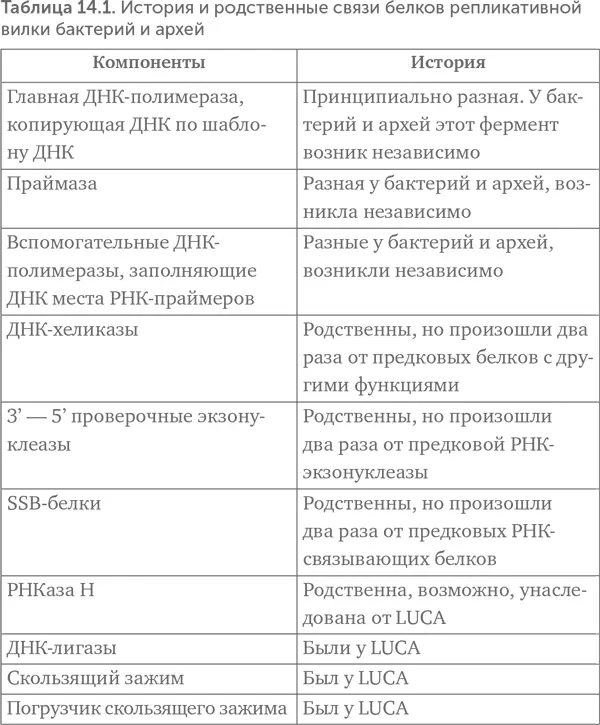
Leipe, Aravind, Koonin (1999) тогда сделали выбор в пользу третьего варианта. По их предположению, LUCA имел гетеродуплексный ДНК-РНК геном, в двухцепочечной форме которого была одна цепь РНК и вторая – ДНК. Такие двойные спирали из разных цепей называются гетеродуплексами. ДНК-РНК-гетеродуплексы прочнее, чем РНК и метил-РНК. Предполагаемый механизм репликации показан на рисунке 14.5 Б. Clamp и Clamp Loader – скользящий зажим и его погрузчик исходно могли помогать обратной транскриптазе не отделяться от копируемой цепи до окончания копирования. Нельзя, впрочем, исключить и другой вариант: у LUCA была репликативная вилка, и в ней работали вместе обратная транскриптаза и ДНК-зависимая РНК-полимераза.
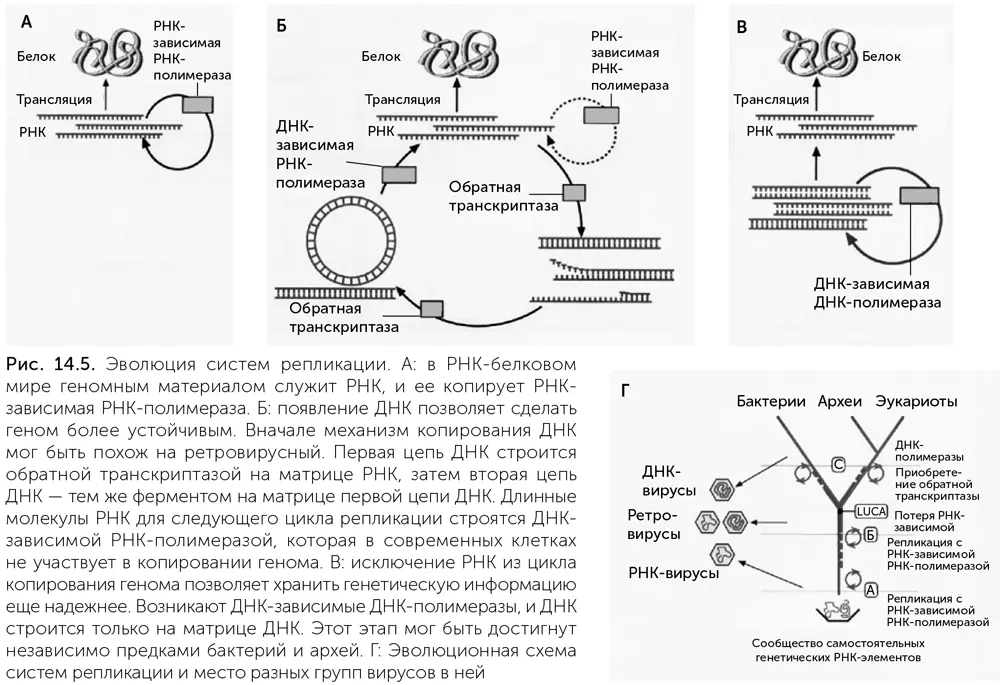
Предположение о гетеродуплексном геноме LUCA легко объясняет, почему бактерии и археи имеют неродственные ДНК-полимеразы. Копировать двухцепочечную ДНК напрямую надежнее, чем через промежуточную стадию РНК, поэтому замена обратной транскриптазы на ДНК-зависимые ДНК-полимеразы была поддержана отбором в линиях бактерий и архей. Это предположение хорошо согласуется и с данными по РНК-полимеразе и ее вспомогательным белкам. Если РНК-полимераза делала молекулы РНК размером в целый геномный фрагмент, то начало и окончание ее работы должны были регулироваться не так, как в современных клетках. Казалось бы, все понятно, но последующие открытия только запутали картину.
Разнообразие и происхождение ДНК-полимераз
В работе Лейпе с соавторами не был учтен тот факт, что все клетки имеют как минимум две ДНК-полимеразы – главную и вспомогательную. Главная полимераза делает основную работу по копированию генома, а вспомогательная заполняет ДНК пустые места на месте удаленных РНК-затравок и поврежденных участков генома. У архей главная и вспомогательная ДНК-полимеразы обычно родственны друг другу и имеют похожие трехмерные структуры, а вот у бактерий две ДНК-полимеразы не похожи ни на архейных «коллег», ни друг на друга. Большинство клеточных полимераз относятся к трем семействам: PolA, PolB и PolC.
К семейству PolB относятся главные и вспомогательные ДНК-полимеразы архей и эукариот, а также подавляющее большинство вирусных ДНК-полимераз. Это семейство имеет трехмерную укладку (фолд) «ладонь и пальцы» (palm-and-fingers), которая характерна и для вирусных обратных транскриптаз, и РНК-зависимых РНК-полимераз. По всей видимости, семейство PolB имеет долгую и богатую историю репликации самых разнообразных геномов (Koonin et al., 2006).
В противоположность им семейство PolC составляют исключительно главные ДНК-полимеразы бактерий. Лишь немногие бактериофаги имеют ДНК-полимеразы семейства PolC, которые они, по всей видимости, недавно позаимствовали у своих хозяев. Трехмерная структура полимераз семейства PolC указывает на их дальнее родство с нуклеотидилтрансферазами – ферментами, достраивающими нуклеотидные цепи без помощи матрицы (Bailey et al., 2006). К ним относятся, например, ССА-трансферазы, участвующие в созревании тРНК, и полиА-трансферазы, достраивающие концы из повторяющихся адениновых нуклеотидов у матричных РНК эукариот.
Наконец, ДНК-полимеразы семейства PolA играют вспомогательную роль у бактерий и копируют геномы некоторых бактериофагов (например, Т7) и митохондрий. Их трехмерная структура отдаленно похожа на укладку «ладонь и пальцы», т. е. они произошли от той же предковой молекулы, что и PolB, но этот предок явно еще не был ДНК-полимеразой.
Кроме этих обычных семейств были обнаружены еще несколько менее распространенных. Семейство PolD найдено у многих архей, но считалось вспомогательным. Однако оказалось, что у Thermococcus геном копирует полимераза PolD, а PolB нужна лишь для ремонта разрывов ДНК (Cubonova et al., 2013). У ряда других архей PolD копирует отстающую цепь, а PolB – лидирующую. Вспомогательные полимеразы семейств PolX и PolY встречаются у отдельных представителей бактерий, архей, эукариот и вирусов. Иначе говоря, ДНК-полимеразы явно возникали в эволюции много раз из ферментов с другими функциями.
Хуже того, оказалось, что функция полимеразы может меняться очень легко. Например, вирус гепатита D копирует свой РНК-геном при помощи ДНК-зависимой РНК-полимеразы хозяина, заставляя ее «читать» РНК вместо ДНК (Macnaughton et al., 2002). У ретровирусов обратная транскриптаза (РНК-зависимая ДНК-полимераза) строит первую цепь двухцепочечного ДНК-генома на матрице РНК, а вторую – на матрице первой цепи ДНК, т. е. тоже читает и РНК, и ДНК. Тип молекулы, который полимераза может «писать», изменить чуть сложнее, но тоже легко. Замена всего двух нуклеотидов превратила ДНК-зависимую ДНК-полимеразу в РНК-полимеразу (Cozens et al., 2012).
В результате к 2015 году картина эволюции систем копирования ДНК окончательно запуталась. В 1999 году Лейпе с коллегами уверенно отвергали возможность замены одной ДНК-полимеразы в клетке на другую, потому что для этого нет очевидных причин. Данные по архейным полимеразам PolD показали, что у архей такие замены точно были, но мы не знаем их причины. Идея Фортера о вирусном происхождении клеточных ДНК-полимераз выглядит красиво и убедительно, но среди известных вирусов есть источники только PolA и PolB. Происхождение PolC от ферментов, строящих цепи РНК без матрицы, объяснить трудно. Происхождение PolD вообще неизвестно. Пока мы не узнали, какой из вариантов сотрудничества PolB и PolD у архей был исходно, трудно сказать что-то определенное про систему репликации LUCA. В рамках гипотезы о LUCA-сообществе возможно, что у него было несколько систем репликации и даже несколько типов генома (РНК, РНК-ДНК гибрид, ДНК) одновременно.
Читать дальшеИнтервал:
Закладка:






